
Richard O. Hynes
Massachusetts Institute of Technology

Erkki Ruoslahti
Sanford Burnham Prebys

Timothy A. Springer
Boston Children’s Hospital/Harvard Medical School
For discoveries concerning the integrins—key mediators of cell-matrix and cell-cell adhesion in physiology and disease
The 2022 Albert Lasker Basic Medical Research Award honors three scientists for discoveries concerning the integrins, key mediators of cell-matrix and cell-cell adhesion in physiology and disease. Independently, Richard O. Hynes (Massachusetts Institute of Technology) and Erkki Ruoslahti (Sanford Burnham Prebys, La Jolla) identified a cell-surface-associated protein that helps affix cells to the surrounding material, the extracellular matrix (ECM). Then the researchers captured a receptor to which this protein binds. In seemingly unrelated studies, Timothy A. Springer (Boston Children’s Hospital/Harvard Medical School) discovered transmembrane proteins that underlie the ability of immune cells to interact with their targets. These initially disparate lines of inquiry would converge and mushroom after the investigators realized that the proteins—now called integrins—belong to the same molecular family. Its members play central roles in an astounding array of processes in embryonic and fully formed organisms, and they offer intervention points for treating myriad diseases.
By around 1900, scientists had ascertained that animal tissues are not random clumps of cells, but rather, highly ordered assemblies. Without adhesion, our bodies would disintegrate, and we now know that cells exist in intimate association with the ECM. Mid-century experiments established the importance of adhesion in development, and the topic converged with cancer as well, yet its molecular basis remained elusive.
Surface explorations, a penetrating idea
By the early 1970s, tumor cells were known to go rogue in the lab as well as the body. Unlike normal cells, they can grow without clinging to culture dishes; furthermore, they ball up and detach instead of spreading out. These unusual properties pointed to the cell surface as a key to some of their aberrant behavior.
Hynes and Ruoslahti, with his collaborator Antti Vaheri (University of Helsinki), independently decided to identify proteins that differentially appear on the surface of normal and malignant cells. The researchers took different tacks but both groups obtained a single abundant protein that coats normal cells and is minimal or absent on cancer cells. Others showed that this protein, which was eventually named fibronectin, fosters the attachment of cells to components of the ECM, and biologists came to appreciate that fibronectin is itself a bona fide member of that material.
Fibronectin addition to cancer cells reversed several of their malignant features. The protein caused them to flatten onto culture dishes, and it restored their internal actin filaments, a major constituent of the molecular skeleton that maintains cell shape. Visualization of actin or fibronectin with fluorescent antibodies demonstrated that these two proteins produce superimposable patterns—with actin inside the cell and fibronectin outside. These observations, from Hynes and others, suggested that some membrane-spanning link connects the molecules. This conjecture raised provocative questions. What was this presumptive molecular hookup and how might it yoke events in the environment to those inside cells?
Dynamic duo
In the meantime, Springer was working in the distant field of immunology. Numerous investigators were racing to track down the T-cell molecule that recognizes infected or malignant cells, which announce themselves by displaying chunks of invaders or suspicious molecules. Certain perplexing observations led Springer to infer that a factor in addition to the canonical T cell receptor was required for this interaction, and he set out to find it. He raised antibodies to surface molecules on cytolytic T cells and tested whether they blocked these cells’ ability to kill their quarry. In 1981, he reported that one such antibody had exactly this effect and thus defined an entity, lymphocyte-function-associated antigen (LFA-1), that executes a vital T-cell task. Further results suggested that LFA-1 helps T cells embrace their targets.
LFA-1 is composed of two distinct proteins, called α and β, and Springer noticed that their sizes correspond to those of another protein, Mac-1, which he had previously identified on a different immune cell, the macrophage. These observations hinted that LFA-1 and Mac-1 might be related. In 1982, he found that each protein’s α and β chain physically associate to form heterodimers—“hetero” because the components differ from each other and “dimer” because there are two. Structural analysis established that the smaller, β subunits of Mac-1 and LFA-1 are close kin or the same, whereas the larger, α subunits differed. Springer soon added a third protein, p150,95, to this growing molecular family. In 1985, he sequenced the first 18 amino acids of the LFA-1 and Mac-1 αs; this fine-tuned examination revealed similarities that implied a relationship in which a single ancestral gene had given rise to multiple αs during evolution.
Springer also discovered that Mac-1 was a previously known receptor that allows macrophages to grasp and engulf microbes and damaged cells. The notion that LFA-1 and Mac-1 perform essential immune functions gained support from observations by Springer and others that a group of individuals with recurrent life-threatening bacterial infections have scant quantities of Mac-1, LFA-1, and p150,95. The underlying condition, Leukocyte Adhesion Deficiency, arises because the common β chain is flawed or underproduced. Consequently, macrophages and other white blood cells cannot appropriately lodge on vessel walls, so they fail to exit the bloodstream and reach infection sites.
Sticky questions, smooth progress
Ruoslahti had been homing in on the part of fibronectin that attaches to cells, a venture that would wind up enabling discovery of the element that ties fibronectin to the cell’s innards. In 1984, he narrowed down this site to an amino-acid stretch that contains an essential arginine-glycine-aspartic acid (RGD in single-letter nomenclature) core. Remarkably, only three of fibronectin’s 1008 amino acids are sufficient to mediate cell attachment.
The following year, he used the molecular tools he had created, including a small peptide that contains RGD, to isolate a protein that binds fibronectin and appeared to be embedded in the cell membrane. Soon afterward, he nabbed a second receptor that tethers a different RGD-containing protein, vitronectin.
Around the same time and within months of each other, Hynes and Ruoslahti reported that they had uncovered yet another receptor that binds RGD-containing proteins, including fibronectin and others involved in clotting. This receptor resides on platelets, cell fragments that slow bleeding and help wounds heal, and it turned out to be a known entity called platelet membrane glycoprotein gpIIb/gpIIIa. David R. Phillips (Gladstone Institute, San Francisco) had established that gpIIb/gpIIIa was, like LFA-1, Mac-1, and p150,95, a heterodimer; he also had demonstrated that platelets from patients with a bleeding disorder called Glanzmann thrombasthenia carry unusually small quantities of gpIIb/gpIIIa. Hynes’s and Ruoslahti’s finding was therefore exciting because it attributed a crucial physiological function to the family of proteins that adhere to the ECM.
Collide and conquer
By then, scientists knew that two components unite to form the fibronectin and the vitronectin receptors. In 1986, Ruoslahti obtained partial sequence of the DNA that encodes these proteins’ α subunits and found that they resemble not only each other, but also the α subunits of LFA-1 and Mac-1.
The likeness did not end there. The scientists would soon find that the β subunits of these proteins were also related. That same year, Hynes isolated DNA that encodes the β subunit of the fibronectin receptor, and a satisfying feature jumped out: It contained a sequence that characterizes transmembrane anchors. These results suggested that the protein was the molecule he had imagined that bridges the cell’s exterior and the actin filaments inside. Hynes named it integrin because it is an integral membrane protein complex that couples the ECM to the cytoskeleton (Figure 1A).
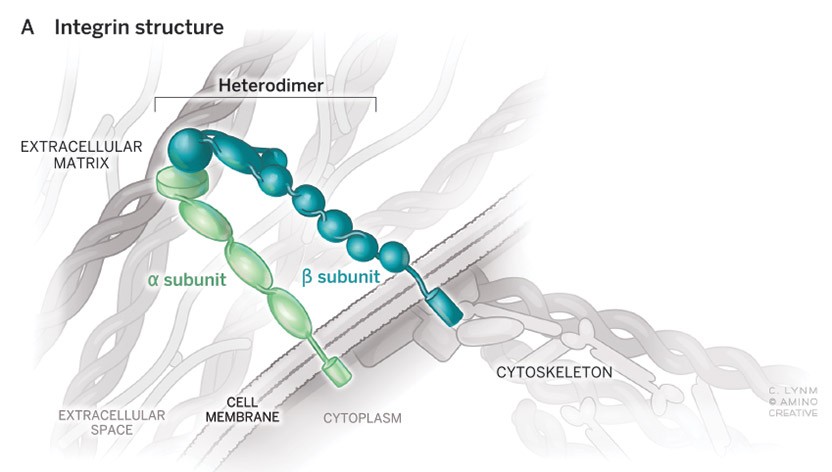
Figure 1A: Pairs that bond
Two protein chains, α and ß, compose the heterodimeric integrins that span the cell membrane. The portion outside affixes to molecules in the extracellular matrix (ECM) or on other cells. Adherence through integrins triggers cytoskeleton assembly and vice versa. Such events can also influence the behavior of other proteins inside the cell. The image shows the matrix-cell type of integrin; a second type mediates cell-cell interactions.
Illustration: Cassio Lynm / © Amino Creative
Within months, Springer reported that a single gene encodes the β chains of LFA-1, Mac-1, and p150,95, and that the integrin β chain was a molecular sibling. He also identified the protein on cells that binds LFA-1 and showed that it defined a new type of partner, as it lacks RGD and belongs to a group of immunoglobulin-like proteins, well known for their roles at the cell surface. The protein that Springer unearthed, intercellular adhesion molecule 1 (ICAM-1), dwells on the endothelial cells that line blood vessels and its residence there explains how LFA-1 foments immune-cell escape from the circulation.
Subsequent work has revealed that humans carry 18 genes for α subunits and eight for β subunits. Various combinations generate 24 known integrins, a name that now applies to all members of the protein family (Figure 1B). Approximately one third of them recognize RGD. These adhesion proteins operate in a plethora of biological enterprises, including the most fundamental processes of cell growth, migration, survival, and proliferation; they likely enabled the evolution of multicellular animals from our unicellular forebears.
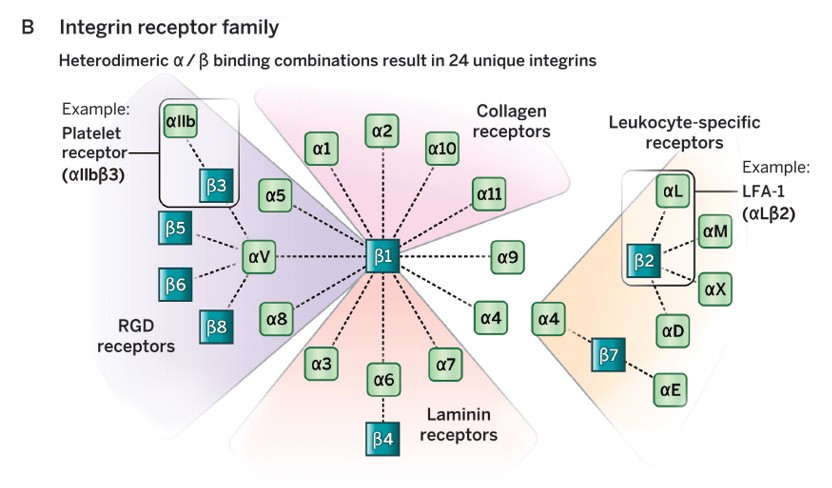
Figure 1B: Families that stick together
Integrin α and ß subunit each belong to a family of proteins with a shared evolutionary ancestor. Individual members of the α and ß families combine in different ways to create 24 mammalian heterodimers that physically associate with a variety of molecules. In the ECM, these binding partners include collagen, laminin, and other RGD-containing proteins such as fibronectin and fibrinogen; on cells, these partners include proteins such as ICAM-1, which dwells on the surface of white blood cells and cells that line blood vessels. The first integrin identified, the platelet receptor, promotes blood clotting. LFA-1 helps immune cells fight infections in numerous ways.
Illustration: Cassio Lynm / © Amino Creative
Cementing discoveries
Scientists have added many layers to our understanding of how integrin-based contact translates into behavioral changes. For example, integrin stimulation spurs a classic molecular event that turns activities on and off: addition of a phosphate group to a particular amino acid on an intracellular protein. This observation, simultaneously reported in 1991 by Hynes and R. L. Juliano (University of North Carolina, Chapel Hill), suggested a means by which integrins might govern physiological affairs and propelled a multitude of studies on their chemical signaling mechanism.
Furthermore, integrins themselves can be activated. Many of these receptors must seize their molecular partners only under particular circumstances. Otherwise, platelets would prompt clotting as they circulate in the bloodstream and T cells would permanently fasten onto cells rather than scan the body for trouble. In 1989, Springer showed that LFA-1 tightens its grip only when the T cell detects another cell that bears foreign proteins and, in response, triggers intracellular events that tune LFA-1’s binding properties. This system allows T cells to circulate until they encounter evidence of infection or other disturbances, at which point the T cell hugs its target.
Figure 2: Fleeting attachments, profound effects
By fostering adherence between cells and the ECM or other cells, integrins play a key role in a tremendous number of essential physiological interactions.
Illustration: Cassio Lynm / © Amino Creative
Burgeoning insights into integrin contributions to health and disease have spawned novel therapeutic strategies for several illnesses. For instance, the eyedrop lifitegrast (Xiida ®) soothes dry eye disease by obstructing LFA-1, one of the integrins that Springer discovered; consequently, white blood cell activity falls, which reduces ocular inflammation. The blockbuster drug vedolizumab (Entyvio ®) also had its origins in Springer’s work, although less directly. This humanized monoclonal antibody selectively thwarts an integrin that promotes immune-cell migration into the gut, thereby quieting ulcerative colitis and Crohn’s disease, both of which result from inflammation of the digestive tract. The agent, which was approved by the FDA for both conditions in 2014, binds an integrin that belongs to the immunoglobulin-like subfamily that Springer first defined. Other compounds, such as tirofiban (Aggrastat ®), aim at cell-ECM rather than cell-cell interactions. This medication inhibits gpIIb/gpIIIa and is used to hamper clotting in certain cardiovascular conditions.
By cracking open the study of integrin-based cell adhesion, Hynes, Ruoslahti, and Springer launched a field that touches countless processes whose mechanisms depend on interchanges among cells and the ECM (Figure 2). Their breakthroughs have illuminated the influence of integrins, which extends throughout the body and the animal world.
by Evelyn Strauss
Selected Publications – Discovery of Integrins
Cell-Matrix Interactions
Ruoslahti, E., Vaheri. A., Kuusela, P., and Linder, E. (1973). Fibroblast surface antigen: a new serum protein. Biochim. Biophys. Acta. 322, 352-358.
Hynes, R.O. (1973). Alteration of cell-surface proteins by viral transformation and by proteolysis. Proc. Natl. Acad. Sci. USA. 70, 3170-3174.
Pierschbacher, M.D., and Ruoslahti, E. (1984). Cell attachment activity of fibronectin can be duplicated by small synthetic fragments of the molecule. Nature. 309, 30-33.
Pytela, R., Pierschbacher, M.D., and Ruoslahti, E. (1985). A 125/115-kDa cell surface receptor specific for vitronectin interacts with the arginine-glycine-aspartic acid adhesion sequence derived from fibronectin. Proc. Natl. Acad. Sci. USA. 82, 5766-5770.
Pytela, R., Pierschbacher, M.D., Ginsberg, M.H., Plow, E.F., and Ruoslahti, E. (1986). Platelet membrane glycoprotein IIb/IIIa: member of a family of Arg-Gly-Asp-specific adhesion receptors. Science. 231, 1559-1562.
Tamkun, J.W., DeSimone, D.W., Fonda, D., Patel, R.S., Buck, C., Horwitz, A.F., and Hynes, R.O. (1986). Structure of integrin, a glycoprotein involved in the transmembrane linkage between fibronectin and actin. Cell. 46, 271-282.
Hynes, R.O. (1987). Integrins: a family of cell surface receptors. Cell. 48, 549-554.
Ruoslahti, E. (1996). RGD and other recognition sequences for integrins. Annu. Rev. Cell Dev. Biol. 12, 697-715.
Hynes, R.O. (2002). Integrins: bidirectional allosteric signaling machines. Cell. 110, 673-687.
Cell-Cell Interactions
Kurzinger, K., Reynolds, T., Germain, R.N., Davignon, D., Martz, E., and Springer, T.A. (1981). A novel lymphocyte function-associated antigen (LFA-1): cellular distribution, quantitative expression, and structure. J. Immunol. 127, 596-600.
Sanchez-Madrid, F., Nagy, J., Robbins, E., Simon, P., and Springer, T.A. (1983). A human leukocyte differentiation antigen family with distinct alpha subunits and a common beta subunit: the lymphocyte function-associated antigen (LFA-1), the C3bi complement receptor (OKM1/Mac-1), and the p150,95 molecule. J. Exp. Med. 158, 1785-1789.
Thompson, W.S., Miller, L.J., Schmalstieg, F.C., Anderson, D.C., and Springer, T.A. (1984). Inherited deficiency of the Mac-1, LFA-1, p150,95 glycoprotein family and its molecular basis. J. Exp. Med. 160, 1901-1905.
Kishimoto, T.K., Lee, A., Roberts, T.M., and Springer, T.A. (1987). Cloning of the beta subunit of the leukocyte adhesion proteins: homology to an extra-cellular matrix receptor defines a novel supergene family. Cell. 48, 681-690.
Makgoba. M.W., Sanders, M.E., Luce, G.E.G., Dustin, M.L., Springer, T.A., Clark, E.A., Mannoni, P., and Shaw, S. (1988). ICAM-1 a ligand for LFA-1-dependent adhesion of B, T and myeloid cells. Nature. 331, 86-88.
Staunton, D.E., Dustin, M.L., and Springer, T.A. (1989). Functional cloning of ICAM-2, a cell adhesion ligand for LFA-1 homologous to ICAM-1. Nature. 339, 61-65.
Springer, T.A. (1990). Adhesion receptors of the immune system. Nature. 346, 425-434.
Lawrence, M.B., and Springer, T.A. (1991). Leukocytes roll on a selectin at physiologic flow rates: distinction from and prerequisite for adhesion through integrins. Cell. 65, 859-873.
Luo, B.-H., Carman, C.V., and Springer, T.A. (2007) Structural basis of integrin regulation and signaling. Annu. Rev. Immunol. 25, 619-647.
Li, J., Yan, J., and Springer, T.A. (2021). Low-affinity integrin states have faster ligand-binding kinetics than the high-affinity state. eLife. 10, 1-22. e73359. doi: 10.7554/eLife.73359.

Building Castles in the Sky and Houses of Cards That Don’t Collapse
The creative process involves two phases—generating new ideas and then focusing on the most tractable and useful ones.

Acceptance remarks
Acceptance remarks, Richard Hynes
Both my parents had scientific backgrounds; I and my three siblings all became scientists, obviously influenced by this environment, and also in my case, aided by an excellent education in a Liverpool city high school and subsequently at Cambridge University Biochemistry in the heyday of the Laboratory of Molecular Biology in the 1960s. I then went to MIT Biology for my PhD; yet another wonderful place to learn how to do science.
I worked on developmental biology at MIT and it’s still one of my scientific loves. I then returned to the UK for a postdoctoral fellowship at the Imperial Cancer Research Fund in London and began to work on the cell biology of cancer. I owe a great deal to all those formative experiences that set me on the road to the research that led to the honour of this Lasker Award—how lucky can one be?
I and my laboratory members at MIT have spent the past 40-plus years exploring the mechanisms of how cells interact with each other and with the extracellular matrix, which is a complex meshwork of proteins to which most cells attach and upon and through which they migrate.
Cell-cell and cell-matrix adhesion organize where cells are positioned in the body, who their neighbors are and how they interact with one another. Integrins are cell-surface receptors that control many of these interactions. They make connections to the extracellular matrix outside and to cytoskeletal machinery inside the cells and act as transducers of signals both into and out of the cells, key to their perception of, and impact upon, their surroundings.
When these connections are altered things can go awry, as in human genetic diseases that affect the adhesion of platelets or white blood cells, leading to various blood disorders, and in cancer when cells do not stay in their proper positions and metastasize to distant parts of the body. Metastasis and the roles of integrins and extracellular matrix proteins are our current areas of research interest—given my background, I think of metastasis as disorganized development.
This has been an exhilarating journey as modern biology has progressed over the past 50 years and powerful new methods of molecular cell biology have made possible experiments that we could not have imagined when we started on this road. The field of cell adhesion and integrins has been a very collegial and collaborative area in which to work. The open and friendly competition has contributed significantly to the scientific advances and will continue to do so in the hands of the many scientists now attracted to this field.
Clinical applications of our understanding of integrins are already having an impact and that will undoubtedly increase given their involvement in so many aspects of embryonic development, physiology and disease.
Acceptance remarks, Erkki Ruoslahti
I am from a small industrial town in southeastern Finland that shared a common border with the Soviet Union, and now with Russia. My father and older brother had engineering degrees from Helsinki Technical University. I decided to try something different and was accepted to study medicine at the University of Helsinki. My plans to become a practicing physician changed after my first year in medical school, when I took a summer job in a hospital bacteriology laboratory and got my first taste of research. This seemed interesting, and I asked if I could work on a research project. The head of the department took me under his wing. He was a great mentor, very keen on supporting young investigators. Several years later, after my postdoc fellowship, I rejoined the same department.
During my studies, we published a lot of papers on serum protein genetics and alpha-fetoprotein in cancer and fetal blood. Several of our papers came out in prominent journals, including Nature, and I thought I was doing quite well. Thinking of where to do a postdoc, I talked at a meeting to an Italian immunologist. He quickly became impatient when I told him about my work on serum protein genetics. He abruptly said, “why don’t you work on something important”—and got up and left. I wasn’t happy about how the interview ended, but having thought about it, I concluded that he had a point and tried keep the advice in mind in choosing projects to work on.
I did my postdoc at Caltech. Rubbing shoulders (and occasional playing tennis) with scientific giants such as Max Delbrück, Ray Owen, and others, was a heady experience for a young researcher from the far northern corner of Europe. At Caltech I was exposed to the ideas that led me to work on what became the recognition system composed of integrins and their ligands, such as fibronectin, in the extracellular matrix.
As my experience illustrates, graduate and/or postdoctoral projects often determine the direction of the rest of one’s scientific career. My advice to young researchers at the beginning of their careers is to think carefully when choosing a mentor and agreeing to work on a project. Ask yourself and older colleagues whose judgement you trust: Is this project worth working on? Will the results be potentially important? If the answer is not a clear yes, switch to do something else.
I am grateful for that long-ago comment about working on something important because it encouraged me to look for significant questions to study and undoubtedly contributed to my receiving this prestigious recognition.
Acceptance remarks, Timothy Springer
It is humbling and a great honor to accept the Lasker Award. I grew up in California with five siblings in a close family that took camping trips every summer. My dad inspired me as resourceful and adventurous. My mom loved and encouraged us. However, I floundered as a freshman at Yale. I studied too little and rebelled against the political establishment running the Vietnam War.
I wanted to do good but did not know how. I started by dropping out to join the Domestic Peace Corps. I served on a Shoshone Reservation in Nevada—living in a little cabin with no electricity, telephone, or running water. The nearest high school was thirty miles away. And in winter, a dirt road over a mountain pass made it impossible to get to. The Shoshones and I wanted to do something about it. So when the governor of Nevada held county road hearings, we made sure to show up and share what the kids in the community were going through. And within a year, that road was being paved.
After a year, I arrived as a transfer student at U.C. Berkeley. I majored in biochemistry and was inspired to learn about protein conformational change from Dan Koshland and protein chemistry from Jack Kirsch. With Jack Strominger, I earned a Ph.D. on biochemistry of cell surface proteins. By an amazing stroke of luck, I did a six-month postdoc in Cambridge England with César Milstein, who had just discovered monoclonal antibodies, for which he’d later win the Nobel Prize.
I then took a faculty position under Baruj Benacerraf at Harvard Medical School. Nothing at the time was known about the molecules that enabled cell-cell recognition in the immune system. I began to discover them by making antibodies that blocked cell-cell interactions, which led to the work I am being recognized for today.
About 40 years have passed since. I am blessed, because there was much to learn about integrin structure and function, and many of the mysteries of how integrins work are only being discovered today. This has required me to learn new techniques including structural biology and single molecule biophysics, and these challenges and the bright students and colleagues with whom I work keep me young. Out of the deep biology my colleagues and I have discovered, new drugs to treat patients have been approved and new companies have been founded. Although I have accepted the challenges of entrepreneurship and philanthropy, I am first and foremost a scientist, and discovery is still what I love most.
