
Katalin Karikó
BioNTech

Drew Weissman
University of Pennsylvania
For the discovery of a new therapeutic technology based on the modification of messenger RNA—enabling rapid development of highly effective Covid-19 vaccines
The 2021 Lasker~DeBakey Clinical Medical Research Award honors two scientists—Katalin Karikó (BioNTech) and Drew Weissman (University of Pennsylvania)—who discovered a new therapeutic technology based on the modification of messenger RNA. This breakthrough enabled rapid development of highly effective Covid-19 vaccines. In addition to providing a tool for quelling a devastating pandemic, the innovation is fueling progress toward treatments and preventives for a range of different illnesses.
In the early 1960s, messenger RNA (mRNA) sprang into the spotlight as the agent that encodes protein, and the possibility of using this nucleic acid for medical purposes began to tickle scientists’ imaginations. mRNA could, in principle, provide a means to turn a person’s cells into factories for any desired protein. Such methodology might replenish essential substances that are in short supply or introduce microbial components as a vaccine. Furthermore, it offered numerous advantages over DNA-based strategies. Unlike DNA, mRNA would not threaten the recipient cell’s genomic integrity because it cannot integrate into the chromosome and interrupt resident genes or wreak other mutational havoc. As an undergraduate in the mid 1970s, Karikó found the promise of mRNA-directed protein replacement irresistible. Since then, she has devoured information and mastered techniques with that enterprise in mind.
For decades, the tantalizing prospects of mRNA therapeutics remained theoretical due to practical impediments. Scientists did not develop ways to synthesize mRNA in a test tube until 1984. After that advance, they still needed to figure out how to deliver it to cells inside animals. Our bodies swarm with enzymes that chew up RNA, and even if the molecule survives the extracellular milieu long enough to encounter a target cell, it then encounters a formidable hurdle: the lipid bilayer. mRNAs are too large to diffuse across this membrane, and they also carry a negative charge, which is rebuffed by the oily barrier.
Although researchers made headway—in the late 1970s, they began to develop lipid-based materials that could package, protect, and transport mRNA to cells and, in 1990, they generated proteins in live rodents by injecting mRNAs into these animals—the overall venture did not gain traction. Concerns about mRNA instability and delivery dominated conventional wisdom, and few investigators had an appetite to tackle these challenges.
A provocative message
But Karikó had committed to her dream and, by the early 1990s, she was a research assistant professor at the University of Pennsylvania, working toward realizing it. During that decade, she made significant findings and persevered despite steady funding rejections and lack of institutional support.
As many journal articles were not yet available online, Karikó photocopied as avidly as she read. In 1997, immunologist Drew Weissman, a new faculty member at Penn, began vying with her for access to the Xerox machine. As the device whirred, the two got talking.
Weissman aimed to make an HIV vaccine. He had not yet tried coaxing cells to construct an immunoreactive HIV protein from an mRNA template, but suddenly an RNA aficionado was offering to make the key reagent for him. He accepted, and the collaborators delivered this mRNA to immature human dendritic cells in culture dishes. In the body, these cells serve as sentinels of the immune system (Ralph Steinman, Lasker Basic Medical Research Award, 2007). Natural encounters with foreign invaders spark transition of the dendritic cells into a mature form that propels T cells into action. Investigators typically instigate this process by adding a stimulating agent. Karikó and Weissman observed that, in addition to prompting abundant manufacture of the HIV protein, the mRNA—on its own—induced a maturation signal. The dendritic cells displayed molecules and behaviors, including competency to galvanize T cells, that characterize the mature state.
Weissman celebrated; Karikó fretted. This attribute of mRNA might invigorate a vaccine, but it would compromise mRNA-based, protein-replacement therapies because mature dendritic cells spit out cytokines, signaling molecules that recruit and provoke other components of the body’s defense system. The resulting inflammation can cause serious harm. As it turned out, some of these immune effects interfere with vaccine applications as well, although the researchers did not know it at the time. Fortunately, they were committed to pursuing Karikó’s quest, so they tried to figure out how to avoid revving up the immune system.
As a first step, they wanted to discern characteristics of the mRNAs that rouse dendritic cells. Toward that end, the investigators tested RNAs from different sources. Bacterial RNA triggered a potent response; total mammalian RNA kindled a mild one. When they homed in further, they found that RNA from human mitochondria, an organelle whose evolutionary origins trace to bacteria, also spurred dendritic cell activation. On the other end of the spectrum, mammalian transfer RNA (tRNA) stirred none. This surprising result provided the crucial clue to Karikó and Weissman’s groundbreaking insight.
Four building blocks compose all RNAs: the nucleosides adenosine, uridine, guanosine, and cytidine. tRNAs are unusual in that they contain a high proportion of so-called modified nucleosides that differ in subtle ways from the standard ones. These variants carry extra chemical groups or have their components attached to one another through different atoms (Figure 1). Karikó and Weissman wondered whether modified nucleosides would render the mRNAs immunologically inert.
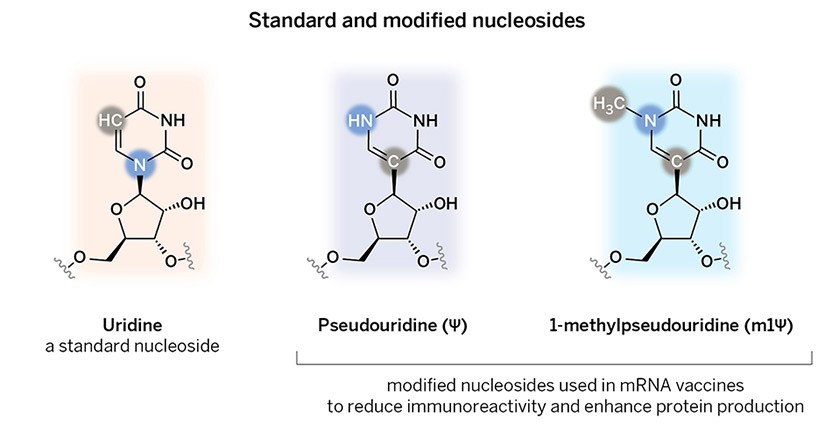
Mighty modifications
RNA that includes the standard nucleoside building block uridine (left) stimulates a strong immune response. Substitution of uridine with pseudouridine (middle) slashes immunoreactivity and increases protein production. Replacement with 1-methylpseudouridine (right) further magnifies both of these beneficial effects. Gray and blue shadings indicate site of modification.
Illustration: Cassio Lynm/© Amino Creative
Here’s looking at U(ridine)
To test this idea, they created a collection of mRNAs, each of which carried a particular modified nucleoside in place of its parent. Then they delivered these mRNAs to dendritic cells. In addition to measuring standard indicators of dendritic cell maturation, the duo assessed activation of two sensors—Toll-Like Receptors (TLR) 7 and 8—that detect single-stranded RNA and churn out cytokines in response. Karikó and Weissman found that substitutions of conventional nucleosides with some of the modified stand-ins softened the inflammatory punch of mRNA because the altered versions failed to activate TLR7 and TLR8, and they published these game-changing data in 2005. Swapping uridine for its common relative, pseudouridine, was especially effective in preventing activation.
The researchers were encouraged by their success at pinpointing the source of immune agitation and devising a way to circumvent it. To be useful for mRNA-based therapies, however, the modified mRNAs also needed to serve as templates for protein production. In 2008, Karikó and Weissman reported that pseudouridine-containing mRNA not only generated functional protein in cell extracts and in cells grown in petri dishes, but boosted manufacture compared to its unmodified, uridine-containing analog. These results held up in mice, and the immunoreactivity observations transferred to the animals as well: pseudouridylation dramatically reduced quantities of interferon-α, a cytokine indicator of inflammation.
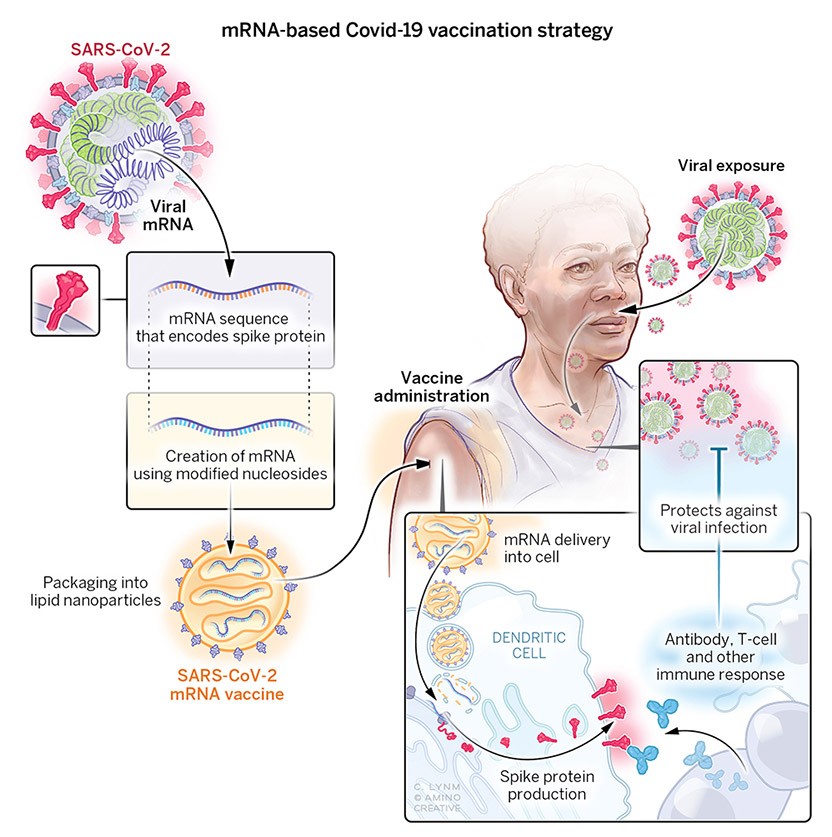
mRNA vaccine scheme
To make a Covid-19 vaccine, scientists sequenced the genetic blueprint for a molecule that plays a key role in infection, the spike protein. Then they synthesized in a test tube a slightly altered version of this mRNA, whose sequence was optimized to encode spike proteins that were known to stimulate an especially effective immune response. The scientists replaced all uridines with 1-methylpseudouridines and encapsulated the resulting mRNA within lipid nanoparticles. After injection, portions of cell membranes pinch off to engulf the particles; the lipid packaging dissolves and releases the mRNA into the resulting sac, where the mRNA encounters TLR7 and TLR8 as it enters dendritic cells. Because the mRNA contains 1-methylpseudouridine rather than its unmodified uridine parent, it does not activate these sensors that would otherwise trigger an immune reaction against the mRNA itself. The modified mRNA is released into the cytoplasm and serves as a template for the cell’s protein-production machinery. The resulting spike protein (red) stimulates production of antibodies (blue) as well as other immune responses, which protect the person upon exposure to SARS-CoV-2.
Illustration: Cassio Lynm/© Amino Creative
Karikó and Weissman braced themselves for a barrage of phone calls. Surely biologists would recognize that the bottleneck on mRNA therapeutics had been shattered. No one rang.
Finally, the significance of their 2005 findings registered on one scientist. In 2010, Derrick Rossi (then at Harvard Medical School) and colleagues used synthetic mRNA that contained modifications to reprogram specialized mammalian cells into pluripotent stem cells. This high-profile paper woke up the scientific community to the prospects and utility of modified mRNA, and Rossi’s findings inspired him to help launch the biotech company Moderna.
In the meantime, Karikó and Weissman pressed on with improvements. Although pseudouridine replacements subdued immunoreactivity, they did not obliterate it, in part because the enzymes that synthesize mRNAs make mistakes that create double-stranded RNAs and other molecules that ignite inflammation. The investigators developed a chromatographic procedure that removes these contaminating molecules, thus enhancing the efficiency of protein production in mammalian cells while eliminating residual safety hazards.
The system was ready for a test run in animals. Karikó and Weissman injected mice with purified mRNA that encodes erythropoietin, a hormone that promotes red blood cell formation. Circulating levels of the substance surged and remained elevated for days, the researchers reported in 2012; furthermore, the erythropoietin fulfilled its normal physiological duties, as measured by an increase in hematocrit levels. The team extended their study to a small number of monkeys, and similarly positive results emerged.
Conquering Covid
By then, it had become clear that Karikó and Weissman’s strategies to curb immunostimulation, which they undertook to enable protein replacement, would benefit vaccine applications as well. Unmodified uridine-containing mRNAs incite production of interferon-α, which thwarts development of T helper cells that are required to generate robust antibody responses and protective immunity.
The pair continued to optimize the method, now working at a distance from one another, as Karikó had moved in 2013 to BioNTech (Mainz, Germany). Shortly thereafter, 1-methylpseudouridine (Figure 1) became available, and it outperformed pseudouridine in taming immunoreactivity and amplifying protein output.
All along, the researchers had used lipid-based packaging to foster cellular uptake of mRNA and guard it from degradation, but every method came with a hitch—toxicity, lability, or some other obstacle to safe and efficient use in humans. Many labs were making substances that would deliver nucleic acid, and each time a new one appeared, Karikó, Weissman, and their colleagues analyzed it. In 2015, Norbert Pardi, a postdoctoral fellow in Weissman’s lab at the time, demonstrated that lipid nanoparticles (LNPs) held enormous potential for clinical use.
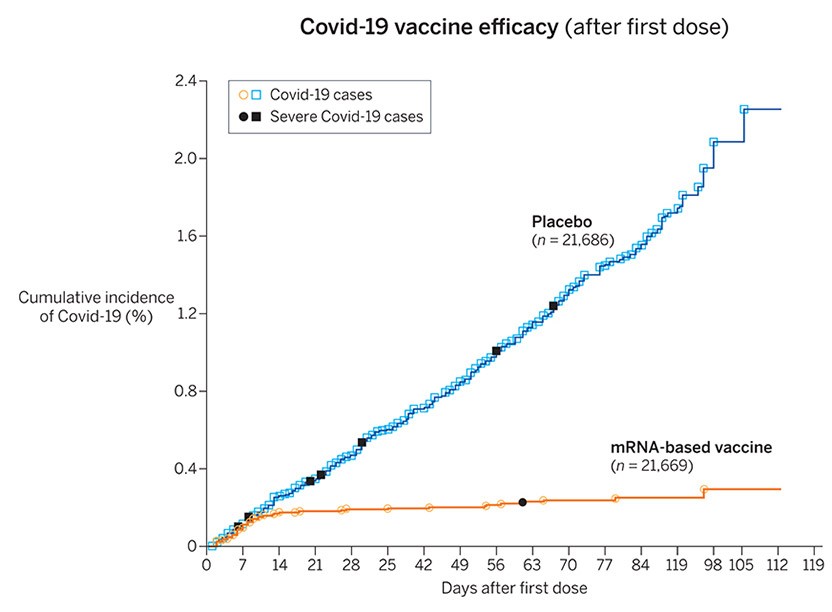
Protection from Covid-19
The BioNTech/Pfizer vaccine protects individuals from Covid-19 infection, beginning about 10 days after inoculation; results for Moderna’s vaccine are similar. Data shown are for the original virus (first detected in Wuhan, China) after the first dose; after two doses, the BioNTech/Pfizer and Moderna vaccines are 94-95% effective. Two doses of these vaccines are also about 90% effective against the Alpha variant (first detected in UK), 60-75% effective against Beta (first detected in South Africa) and Gamma (first detected in Brazil) variants, and 85% effective against the Delta variant (first detected in India). Each symbol denotes the total number of Covid-19 cases, defined by symptoms and presence of virus; filled symbols denote severe cases. The x axis shows time post injection with vaccine or placebo.
Reprinted with permission from Massachusetts Medical Society. Polack FP et al., N Engl J Med. 2020;383:2603-2615.
Less than two years later, Pardi, Weissman, and Karikó harnessed the elements they had developed to design a candidate vaccine against Zika virus. A single inoculation provided rapid and lasting immunity in mice and monkeys. Thus, when the SARS-CoV-2 pandemic hit, the pieces were in place (Figure 2). In January 2020, when the sequence of the new coronavirus became available, BioNTech/Pfizer and Moderna used it to make vaccines in record time that are 95% effective in preventing symptomatic infection from the original viral variant (Figure 3). To date, hundreds of millions of people throughout the world have been injected with one of these two vaccines, both of which have proved to be remarkably safe.
Many groups are pursuing modified mRNA-based vaccines for a plethora of diseases, and the scheme is proving its versatility. Researchers are developing it to supply therapeutic proteins that combat a wide array of illnesses, including cancers, infectious diseases, and autoimmune disorders. They are trying to fix gene defects in sickle cell anemia, harness the CRISPR-Cas9 editing system to destroy other troublemaking DNA, and much more. Karikó and Weissman’s relentless efforts have already saved countless human lives from the ravages of Covid-19, and their pivotal discoveries promise to revolutionize medical science well beyond the current microbial scourge.
by Evelyn Strauss
Selected Publications of Katalin Karikó and Drew Weissman
Ni, H., Capodici, J., Cannon, G., Communi, D., Boeynaems, J.-M., Karikó, K., and Weissman, D. (2002). Extracellular mRNA induces dendritic cell activation by stimulating tumor necrosis factor-a secretion and signaling through a nucleotide receptor. J. Biol Chem. 277, 12689-12696.
Karikó, K., Buckstein, M., Ni, H., and Weissman, D. (2005). Suppression of RNA recognition by toll-like receptors: the impact of nucleoside modification and the evolutionary origin of RNA. Immunity. 23, 165-175.
Karikó, K., Muramatsu, H., Welsh, F.A., Ludwig, J., Kato, H, Akiro, S., and Weissman, D. (2008). Incorporation of pseudouridine into mRNA yields superior nonimmunogenic vector with increased translational capacity and biological stability. Mol. Ther. 16, 1833-1840.
Karikó, K., Muramatsu, H., Ludwig, J., and Weissman, D. (2011). Generating the optimal mRNA for therapy: HPLC purification eliminates immune activation and improves translation of nucleoside-modified, protein-encoding mRNA. Nucleic Acids Res. doi: 10.1093/nar/gkr695
Karikó, K., Muramatsu, H., Keller, J.M., and Weissman, D. (2012). Increased erythropoiesis in mice injected with submicrogram quantities of pseudouridine-containing mRNA encoding erythropoietin. Mol. Ther. 20, 948-953.
Pardi, N., Tuyishime, S., Muramatsu, H., Karikó, K., Mui, B.L., Tam, Y.K., Madden, T.D., Hope, M.J., and Weissman, D. (2015). Expression kinetics of nucleoside-modified mRNA delivered in lipid nanoparticles to mice by various routes. J. Control. Release. 217, 345-351.
Pardi, N., Hogan, M.J., Pelc, R.S., Muramatsu, H., Andersen, H., DeMaso, C.R., Dowd, K.A., Sutherland, L.L., Scearce, R.M., Parks, R., Wagner, W., Granados, A., Greenhouse, J., Walker, M., Willis, E., Yu, J.-S., McGee, C.E., Sempowski, G.D., Mui, B.L., Tam, Y.K., Huang Y.-J., Vanlandingham, D., Holmes, V.M., Balachandran, H., Sahu, S., Lifton, M., Higgs, S., Hensley, S.E., Madden, T.D., Hope, M.J, Karikó, K., Santra, S., Graham, B.S., Lewis, M.G., Pierson, T.C., Haynes, B.F., and Weissman, D. (2017). Zika virus protection by a single low-dose nucleoside-modified mRNA vaccination. Nature. 9, 248-251.
Perspectives on mRNA Vaccines
Sahin, U., Karikó, K., and Türeci, O. (2014). mRNA-based therapeutics–developing a new class of drugs. Nature Rev. 13, 759-780.
Pardi, N., Hogan, M.J., Porter, F.W., and Weissman, D. (2018). mRNA vaccines–a new era in vaccinology. Nature Rev. 17, 281-279.
Rubin, E.J., and Longo, D.L. (2020). SARS-CoV-2 vaccination–an ounce (actually, much less) of prevention. N. Engl. J. Med. 383, 2677-2678.
Haynes, B.F. (2021). A new vaccine to battle Covid-19. N. Engl. J. Med. 384, 470-471.

The Surprise Element: A Distinctive Characteristic of Creative Scientists, Artists, and Comedians
Truly creative works of science and art produce unexpected and surprising results—just like the punch line of a good joke that generates an unfamiliar twist on a familiar idea.
Acceptance remarks
Acceptance remarks, Katalin Karikó
I grew up in a small Hungarian town with a population of ten thousand. As a little girl I liked to climb up trees to check out the bird nests and watch the neighbor’s cow give birth. I was curious. My science teachers made sure that I stayed curious for life and encouraged me to keep learning.
Though I had never actually seen a scientist, I decided that’s what I will do. Now today receiving the Lasker award, I have been able to reflect on my humble beginnings and the long winding road on which I have arrived.
In high school my teacher handed me a book called The Stress of Life in which Hans Selye wrote that “adopting the right attitude can convert a negative stress into a positive one.” This book was my guide through the years as I continued the journey of becoming a scientist.
I remembered Selye’s words when I experienced failures–as a student, or in the lab–so I adopted the right attitude, searched for ways to improve myself, to work harder, be more creative and perform better. I always try to focus on things that I can do and not waste any time on things that I cannot.
I began my work with RNA as a graduate student in 1978 and have been passionate about this fragile molecule ever since. I learned the ways that RNA can be made and modified, how it degrades or translates into proteins.
I love the enigmatic nature of RNA-related phenomena, and enjoy revisiting experimental outcomes that I couldn’t explain decades earlier. I am always delighted when scientific puzzles that I tried to decode myself are ultimately solved by others.
My greatest joys in science have been the small and large victories in the lab. Even if an experiment fails, it teaches us something that we could build upon.
Together with my colleagues we inched towards a solution, building upon discoveries of scientists who came before us, and created the optimal RNA suitable for therapy. Never in a million years would I have imagined that it would be used to create a vaccine to combat a global pandemic.
While I am accepting my Lasker Award, I think about all those hard-working fellow scientists who are–just like me–passionate about their work and immensely contributed to the success of the Covid-19 mRNA vaccine.
I also think about all of the young girls who may become inspired by my story and want to become scientists. To them I say: stay curious, adopt the right attitude and stay on the track no matter how long and winding that road may be.
Acceptance remarks, Drew Weissman
Those who know me, know that I’m more at home in my lab than I am in the spotlight. And no offense intended, but I would much prefer to be there than in front of a camera right now…as you may be able to tell.
My wife, Mary Ellen, and children Rachel and Allison, are often frustrated by my disinterest in celebrating my own successes. When research indicated that mRNA vaccines surpassed 90 percent effectiveness at preventing Covid-19 and almost 100 percent at preventing hospitalizations from it, my family wanted to do something special. Once we were fully vaccinated, I said, “Alright, if you really want to celebrate, I guess we can go have a nice dinner.” I just wanted to get back to the lab.
I am, however, truly grateful that work I did has helped the world. That’s the hope of every physician-scientist. And I am honored to be the recipient of a Lasker Award and to be counted among a remarkable group of recipients, which includes my long-time scientific partner, Katalin Karikó.
At Penn, Dr. Kariko and I investigated mRNA as a medical intervention. We were both excited by the promising possibilities of an area of medicine we did not feel had been sufficiently explored. And after all, we were at the University of Pennsylvania, an institution of “firsts”–the nation’s first medical school and a place where breakthroughs happen.
So, we conducted experiments. Each spark of something interesting, whether a finding we expected–or even more exciting, the ones we didn’t–motivated us to continue. Together we designed and conducted experiments, often trading emails into the early hours–unable to wait until the next day to share particular findings or ideas. We just kept at it.
Although the investigations that we began over two decades ago have culminated in significant discoveries and a vaccine against a pandemic-causing virus, the work continues. I’m thrilled to say that my lab is pursuing new mRNA vaccines to guard against a host of infectious diseases, like influenza and HIV, and even a pan-coronavirus vaccine that could offer protection against all viruses in the coronavirus category.
Equally important to discovery is ensuring that everyone is able to realize the benefits. We are collaborating with researchers around the world to help them create their own mRNA vaccines for Covid-19 to increase the global supply for people in low- and middle-income countries. And we are taking opportunities to educate people on the science behind this often-misunderstood vaccine.
Scientists know the work is never over because, as much as it’s black and white when looking at data on the page, the implementation and utilization of discovery is open-ended. The work is never over because of the need for scientific breakthroughs: not just for the current pandemic, but for autoimmune diseases, neurologic diseases, and rehabilitation after trauma, and other infectious diseases. Making the discovery and answering your own curiosity is thrilling but making an impact on human beings is what excites me the most.
Today, as I take a second to appreciate this good fortune, that our work has helped millions of people around the world, I also want to recognize the scientists who are “at home” in their own labs right now, fueling the engine of progress, doing the work that may lead to something amazing. I’ll join you back home in my lab in the morning…or if I get antsy, maybe in a few hours. Together, we’ll see what the future holds.
