
Richard H. Scheller
Genentech

Thomas C. Südhof
Stanford University School of Medicine
The 2013 Albert Lasker Basic Medical Research Award honors two scientists for their discoveries concerning rapid neurotransmitter release, a process that underlies all of the brain’s activities. Richard H. Scheller (Genentech) and Thomas C. Südhof (Stanford University School of Medicine) identified and isolated many of this reaction’s key elements, unraveled central aspects of its fundamental mechanism, and deciphered how cells govern it with extreme precision. These advances have provided a molecular framework for understanding some of the most devastating disorders that afflict humans as well as normal functions such as learning and memory.
The billions of nerve cells in our brains allow us to savor chocolate, whack a baseball, and imagine traveling at the speed of light. Their exploits tell our hearts to quicken and make us feel as if those same hearts are breaking. Their messages give us eureka moments — and let us jump out of the bathtub in response.
In the 1950s, the late Bernard Katz figured out that cells eject neurotransmitters in fixed amounts. Electron-micrographic studies by others illuminated how. Balloon-like containers — vesicles — each hold set quantities of the chemicals. Calcium incites these lipid-bound sacs to fuse with the membrane that encases the cell, and their contents spill out (see Figure).
Neurotransmission occurs astonishingly quickly — fast enough for a person to pull a hand off a hot burner or dodge an attacking mountain lion. Calcium entry can spark the release of neurotransmitter packages in less than a millisecond.
Although this general scheme had emerged by the time Südhof and Scheller began their work, its molecular details eluded scientists. No one knew what drives the vesicles to fuse with the cell membrane or how calcium provokes that event.
Actors at the membrane
In the late 1980s, Scheller and Südhof set out independently to unveil the molecular underpinnings of the process. Reasoning that nerve terminals would hold the reaction’s components, the scientists decided to purify and characterize proteins at that site. Focusing first on vesicles, Scheller, then at Stanford University School of Medicine, discovered what would turn out to be an essential piece of the fusion apparatus: vesicle-associated membrane protein (VAMP), which he obtained from the electric organ of a marine ray. The following year, Südhof, then at the University of Texas Southwestern Medical Center in Dallas, isolated the same protein from rat brain and called it synaptobrevin.
Südhof next purified and studied another vesicle protein, synaptotagmin. When calcium is present, this protein binds to phospholipids, major constituents of membranes. Scheller established that synaptotagmin clutches a brain protein that he named syntaxin. In contrast to vesicle-dwelling synaptotagmin and VAMP/synaptobrevin, syntaxin concentrates in the cell membrane — at spots where neurotransmitters are released from nerve cells.
Independently, James Rothman (Yale University) (Lasker Basic Medical Research Award, 2002) had been exploring how substances are ferried from one place to another inside cells. In that process, too, transport vesicles deliver their contents by merging their membranes with those of the target compartment. Rothman had proposed that one of the proteins necessary for his experimental system and for fusion in live yeast cells, NSF, attaches to the membrane through a second protein and its as-yet-unidentified collaborators. He set out to unearth these hypothetical proteins, which he called SNAREs.
To nab them, Rothman sought proteins in rat brains that adhere to NSF through its partner. Three proteins stuck: VAMP/synaptobrevin, syntaxin, and SNAP-25, a protein reported previously to reside on cell membranes of nerve termini.
Two separate lines of inquiry thus pointed at the same three molecules. The results suggested that they promote neurotransmitter release by fostering fusion, but provided only indirect evidence. Meanwhile, a clear link to their physiological function had materialized from a different direction. Scientists had long known that certain bacterial toxins thwart neurotransmission, and in 1992, Cesare Montecucco (University of Padua) showed that botulinum and tetanus toxins block neurotransmitter release by clipping off a chunk of VAMP/synaptobrevin. Subsequent work by Montecucco, Südhof, and Reinhard Jahn (Max Planck Institute, G ttingen) demonstrated that additional neurotoxins attack syntaxin and SNAP-25.
These observations established that the three proteins are vital for neurotransmission. The mechanism of fusion was murky, however, as was the way in which calcium triggers the process.
Findings fuse
The previous work had shown that VAMP/synaptobrevin, syntaxin, and SNAP-25 bind to the NSF conglomeration, but not whether they do so alone or together. In 1993, Scheller, in collaboration with Rothman, found that VAMP/synaptobrevin, syntaxin, and SNAP-25 associate stably with one another to form the so-called ‘SNARE complex’. The team also showed that NSF rips apart this assemblage. Scientists later realized that this phenomenon helps recycle the molecular machinery.
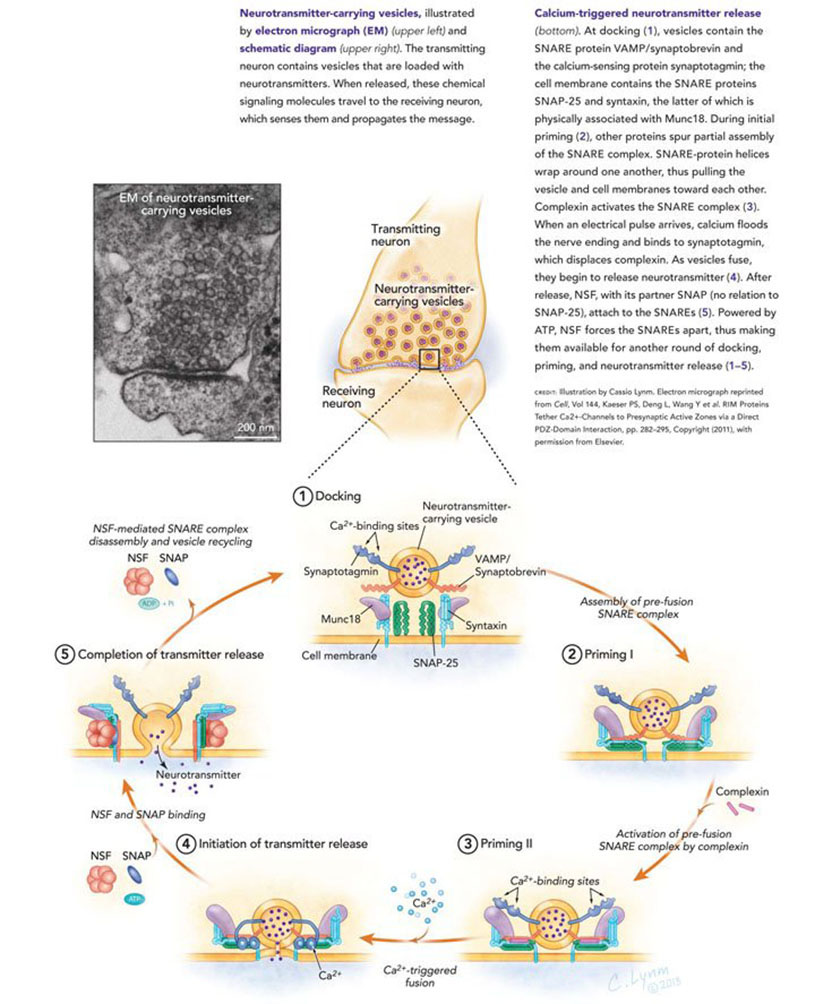
In the meantime, researchers were defining the precise interactions among these proteins and discerning how the associations might instigate fusion (see Figure for current model). Scheller found, for instance, that VAMP/synaptobrevin on vesicles clasps plasma membrane-bound syntaxin, and that SNAP-25 helps them pair up. Work by Scheller, Jahn, John Heuser (Washington University School of Medicine, St. Louis), and others advanced the concept that VAMP/synaptobrevin and syntaxin form coils that wrap around each other along their lengths, thus drawing together the membranes in which they’re embedded. Axel Brunger (Yale University) and Jahn confirmed and extended these ideas when they reported the high-resolution structure of VAMP/synaptobrevin, syntaxin, and SNAP-25 in 1998. The results supported a scenario in which the proteins zipper together, eventually forcing fusion (see Figure).
Control mechanisms calcify
These and other observations were fleshing out the basic mechanism of fusion, but they also highlighted crucial open questions. Uncontrolled, the reaction would result in rampant and constant neurotransmitter release. Although scientists knew that the system does not launch until calcium arrives, the details remained obscure. It was clear, however, that the extremely short time between calcium influx and neurotransmitter discharge would not permit assembly of a multi-protein machine. The device must lie on the brink of its fusion-competent state, waiting for calcium to push it over the edge, presumably by a protein that senses this ion.
Südhof’s original observations on synaptotagmin tantalized him because they indicated that the protein performs calcium-dependent activities. Perhaps, he speculated, calcium prompts it to facilitate fusion. In 1993, he showed that two regions of synaptotagmin bind calcium, and this property allows it to efficiently grasp phospholipids. These results bolstered the protein’s candidacy as the calcium sensor.
In 1994, he generated mice that lack operational synaptotagmin. Although the animals died soon after birth, neurons from their embryos could be studied. Südhof found that their core fusion machinery remained functional, but unable to respond to calcium.
In an elegant set of experiments, Südhof ruled out the possibility that loss of synaptotagmin impairs calcium entry into the cell or hinders some other event peripheral to calcium perception. He generated a series of mice, each of which carried a synaptotagmin with altered calcium-binding affinity. Synaptotagmin’s affinity for the ion correlated with the calcium sensitivity of neurotransmission. Increasing calcium avidity, for instance, decreased by approximately the same amount how much calcium was needed for neurotransmitter release. These results confirmed the notion that synaptotagmin functions as the calcium sensor.
Meanwhile, another layer of regulation was surfacing. Südhof had discovered a protein, complexin, that strongly adheres to the VAMP/synaptobrevin, syntaxin, and SNAP-25 bundle. Subsequent work established that it plays an essential role in calcium-regulated neurotransmitter release. Complexin holds the partially zippered SNARE complex in a form that is poised to trigger fusion, yet inactive until the crucial next step: Calcium binds to synaptotagmin and spurs it to displace complexin, which instantly drives the reaction (see Figure).
Medical implications
Communication within the brain influences how we think and who we are. Defects in the process contribute to schizophrenia, depression, bipolar disorder, and many other pathological conditions. Studies of these illnesses have not yet indicted misbehaving components of the fusion complex itself, but scientists are beginning to uncover connections between this equipment and serious diseases. For example, α-synuclein, a protein that has been linked to Parkinson disease and other neurodegenerative disorders, helps assemble the fusion apparatus and protects animals from age-related neurological problems. Flaws in Munc18, another protein that Südhof and Scheller implicated in neurotransmitter release and that is indispensable for it, have been associated with Ohtahara syndrome, a severe epileptic disorder that strikes during infancy and causes seizures and mental retardation.
By systematically exposing and analyzing the proteins involved in neurotransmitter release, Südhof and Scheller have transformed our description of the process from a rough outline to a series of nuanced molecular transactions. Their work has revealed the elaborate orchestrations that lie at the crux of our most simple and sophisticated neurobiological activities.
by Evelyn Strauss
Key publications of Richard H. Scheller
Trimble, W.S., Cowan, D.M., and Scheller, R.H. (1988). VAMP-1: A synaptic vesicle-associated integral membrane protein. Proc. Natl. Acad. Sci. USA. 85, 4538-4542.
Bennett, M.K., Calakos, N., and Scheller, R.H. (1992). Syntaxin: A synaptic protein implicated in docking of synaptic vesicles at presynaptic active zones. Science. 257, 255-259.
Söllner, T., Bennett, M.K., Whiteheart, S.W., Scheller, R.H., and Rothman, J.E. (1993). A protein assembly-disassembly pathway in vitro that may correspond to sequential steps of synaptic vesicle docking, activation, and fusion. Cell. 75, 409–418.
Calakos, N., Bennett, M.K., Peterson, K.E., and Scheller, R.H. (1994). Protein-protein interactions contributing to the specificity of intracellular vesicular trafficking. Science. 263, 1146-1149.
Lin, R.C. and Scheller, R.H. (1997). Structural organization of the synaptic exocytosis core complex. Neuron. 19, 1087-1094.
Chen, Y.A., Scales, S.J., Patel, S.M., Doung, Y.-C., and Scheller, R.H. (1999). SNARE complex formation is triggered by Ca2+ and drives membrane fusion. Cell. 97, 165-174.
Key publications of Thomas C. Südhof
Perin, M.S., Fried, V.A., Mignery, G.A., Jahn, R., and Südhof, T.C. (1990). Phospholipid binding by a synaptic vesicle protein homologous to the regulatory region of protein kinase C. Nature. 345, 260-263.
Brose, N., Petrenko, A.G., Südhof, T.C., and Jahn, R. (1992). Synaptotagmin: A calcium sensor on the synaptic vesicle surface. Science. 256, 1021-1025.
McMahon, H.T., Missler, M., Li, C., and Südhof, T.C. (1995). Complexins: Cytosolic proteins that regulate SNAP receptor function. Cell. 83, 111-119.
Fernández-Chacón, R., Königstorfer, A., Gerber, S.H., Garcia, J., Matos, M.F., Stevens, C.F., Brose, N., Rizo, J. Rosenmund, C., and Südhof, T.C. (2001). Synaptotagmin I functions as a calcium regulator of release probability. Nature. 410, 41-49.
Rhee, J.-S., Li, L., Shin, O.-H., Rah, J.-C., Rizo, J., Südhof, T.C., and Rosenmund, C. (2005). Augmenting neurotransmitter release by enhancing the apparent Ca2+ affinity of synaptotagmin 1. Proc. Natl. Acad. Sci. USA. 102, 18664–18669.
Tang, J., Maximov, A., Shin, O.-H., Dai, H., Rizo, J., and Südhof, T.C. (2006). A complexin/synaptotagmin-1 switch controls fast synaptic vesicle exocytosis. Cell. 126, 1175-1187.
Review articles by Scheller and Südhof on neurotransmitter release
Bennett, M.K. and Scheller, R.H. (1993). The molecular machinery for secretion is conserved from yeast to neurons. Proc. Natl. Acad. Sci. USA. 90: 2559-2563.
Südhof, T.C. (1995). The synaptic vesicle cycle: A cascade of protein-protein interactions. Nature. 375, 645-653.
Lin, R.C. and Scheller, R.H. (2000). Mechanisms of synaptic vesicle exocytosis. Annu. Rev. Cell Dev. Biol. 16, 19-49.
Südhof, T.C. (2012). Calcium control of neurotransmitter release. Cold Spring Harb. Perspect. Biol. 4:a011353, 1-15.
Award presentation by Eric Kandel
 We are who we are because of our brain and its ability to acquire and store new information. This ability derives from the remarkable capabilities of the 100 billion signaling units in the brain called nerve cells — or neurons — and in particular from the ability of these nerve cells to communicate with one another through specialized contact points called synapses. A synapse has three components: a pre-synaptic terminal contributed by the sending neuron, a post-synaptic receptive component contributed by the receiving cell and a synaptic cleft — the space between the pre- and post-synaptic components. Synapses operate by means of a chemical called a neurotransmitter that is released from the presynaptic terminal by a process involving the influx of Ca2+, whose molecular bases our two Lasker Basic Science Prize winners, Richard Scheller and Thomas Sudhof, have pioneered in defining.
We are who we are because of our brain and its ability to acquire and store new information. This ability derives from the remarkable capabilities of the 100 billion signaling units in the brain called nerve cells — or neurons — and in particular from the ability of these nerve cells to communicate with one another through specialized contact points called synapses. A synapse has three components: a pre-synaptic terminal contributed by the sending neuron, a post-synaptic receptive component contributed by the receiving cell and a synaptic cleft — the space between the pre- and post-synaptic components. Synapses operate by means of a chemical called a neurotransmitter that is released from the presynaptic terminal by a process involving the influx of Ca2+, whose molecular bases our two Lasker Basic Science Prize winners, Richard Scheller and Thomas Sudhof, have pioneered in defining.
Acceptance remarks

Acceptance remarks, 2013 Lasker Awards Ceremony
I would like to take a couple of minutes to reflect on the progress made in cellular and molecular neuroscience and to then comment on what I see as some of the biggest problems facing the field, particularly in the area of human health.
I went back and looked at the proceedings of the first National Institute of Mental Health Conference on Molecular Neurobiology that was held in Santa Barbara, California, 25 years ago. Steve Zalcman and I organized the conference, and I’m happy to note that Steve is in the audience today. The problems we were attempting to solve were not new but we were armed with a new set of molecular, cellular, and genetic techniques that gave us hope for unprecedented progress. Interestingly, the field was relatively small at the time. Most of the important people in the field fit into a medium sized room, all spoke during the three-day conference, and many are here today.
My first observation is that the breadth of this field has grown immensely such that topics represented 25 years ago by individual speakers would now require individual meetings.
Second, while it may not feel that way day to day in the lab, in my opinion progress has been meteoric. This is confirmed by the fact that 5 of the people who attended that first conference have since won the Nobel Prize and others have received the Lasker, Kavli, Gardner, Gruber, and other prizes. We have gained tremendous insight, or essentially solved, many important problems in the field. I do not have time to go over that progress in detail, which in any case does not need to be recounted for this learned audience.
Third, when I think about how this progress will be applied to the important medical problems facing mankind, I am both excited and a bit concerned.
I believe that we have a number of insights that provide those of us interested in therapeutic intervention reason to be optimistic about our progress working with diseases of neuronal degeneration, such as Alzheimer’s, ALS, and Parkinson’s. The genetics of these conditions has uncovered pathways that are likely important in causing the diseases, at least in certain subsets of patients. The ability to dissect the pathways with molecular and cellular techniques has pointed us toward mechanism and in some cases suggested ways one might intervene with therapeutics.
In contrast, I am much less optimistic about our ability to make rapid medical progress in psychiatric disorders such as depression, schizophrenia, and autism. These diseases manifest themselves through the immense complexity of neural circuitry and are perhaps neurodevelopmental disorders. Whatever their origin, we need a much greater understanding of the brain before we will have a mechanistic understanding of the cause of these psychiatric disorders which will then guide us towards effective treatments.
What do we do about this? I have no answers except a trivial solution. Keep working hard. I believe that if the progress in the next 25 years is as tremendous as the last 25 years we will have insights into these devastating diseases that will provide major benefits for patients and society.
Finally, conducting this research is expensive. Unfortunately, over all NIH funding, corrected for inflation, peaked about a decade ago and has even more dramatically decreased on a per laboratory basis. I know many of us hope this trend will be reversed so the promise of understanding the brain and its devastating diseases can be realized.

Acceptance remarks, 2013 Lasker Awards Ceremony
I grew up in an academic household with a strong religious bent. True to my name and nature, from early on I doubted the tenets of my parents’ beliefs — I probably was not easy to have around (hasn’t changed …). What my upbringing instilled in me was a desire to question what is actually true. My greatest pleasure has always been to discover facts, to figure out how something works, to identify the relationships and connections that explain an observation. I feel particularly honored by this award because the Lasker Award recognizes long-lived advances of medical knowledge that produce true insights, rather than some short-lived intriguing “discovery” that appears on the front page of the New York Times.
I studied medicine because I wanted to do something useful, but I soon started to do science on the side. Initially, I was a biophysicist and biochemist who studied organelles and proteins in Victor Whittaker’s laboratory in Göttingen. Whittaker was the scientist who first isolated synaptic vesicles in the 1960s. Shortly after finishing medical school in 1982, I became a postdoctoral fellow with Mike Brown and Joe Goldstein at UT Southwestern in Dallas, where I learned to be a molecular cell biologist and studied the LDL receptor gene and cholesterol regulation. After finishing my post-doc, I merged my earlier love of synaptic vesicles with my new knowledge of how to purify and clone membrane proteins, with the ultimate goal of pursuing something entirely new — to identify all the membrane proteins that make up a synaptic vesicle, and to figure out how these vesicle proteins mediate release of neurotransmitters.
Synaptic vesicles are chock-full with neurotransmitters and release them by fusing with the presynaptic plasma membrane. Neurotransmitter release underlies all communication of neurons with each other at synapses. When I began this work in 1986, none of the proteins of synaptic vesicles had been identified. Nothing was known about neurotransmitter release except that it was triggered by calcium. Whittaker had introduced me to the subject of synaptic vesicles and neurotransmitter release, but it was Joe Goldstein and Mike Brown who had taught me how to actually approach this subject, and how to study how a synapse works.
Over the last 25 years, I mutated not only mouse genes, but changed myself, morphing from a molecular biochemist into a mouse geneticist into an electrophysiologist into a behavioral neuroscientist. These changes gave me the pleasure of always learning something new about how synaptic vesicles and synapses work, although these changes unfortunately have not improved my looks.
The 1990ies were an amazingly thrilling time! During this time, we together with our long-time collaborators Reinhard Jahn and Jose Rizo, and in parallel with Richard Scheller and others, discovered the role of SNARE and SM proteins in synaptic vesicle fusion. We identified synaptotagmins as calcium-sensors in neurotransmitter release, showed how synapses are organized by active zone proteins, and described the first synaptic cell-adhesion molecules that guide synapse formation. These discoveries and conceptual advances were the beginning of an understanding of synapses, which has now become generally recognized. During the course of our work, the studies of Jim Rothman, who received a Lasker Award in 2002 for his work on the cell biology of vesicle fusion, expanded our thinking and injected great enthusiasm into the rapidly growing field of neurotransmitter release.
I thank the Lasker Foundation — I thank you all for giving me this award, and I hope to live up to it!

Acceptance remarks, 2013 Lasker Awards Ceremony
Development of the modern cochlear implant was a worldwide effort involving many scientists, engineers, physician-scientists, and research subjects. The success of this effort is an outstanding example of the power of collaborations between the public and private sectors and also the informed support by the NIH of applied as well as basic research.
I am proud to stand before you today as a representative of the worldwide effort, and I am especially proud to stand with Graeme Clark and Ingeborg Hochmair, who are two of my heroes and the foremost living pioneers in our field.
Although the present cochlear implants are truly wonderful, room still exists for improvements. A variability in outcomes remains, and even the top-performing patients experience difficulties in understanding speech in adverse acoustic environments such as noisy restaurants or workplaces. In addition, reception of sounds more complex than speech — such as music — is less than satisfying for most patients. Research is underway to narrow or even eliminate these gaps between prosthetic and fully functional hearing, and to narrow the range of outcomes such that all patients will achieve high levels of performance. Many promising possibilities are being pursued by extraordinarily talented investigators, and I am completely confident that further improvements will be made.
An even more important challenge — in my mind — is to make the highly effective technology we have today available to all persons who could benefit from it. Thus far, about 320 thousand persons have received a cochlear implant in one or both ears. But various estimates indicate that as many as 25 million persons worldwide could benefit from a cochlear implant. That means that only about 1 or 2 percent of the population who could benefit actually have received a cochlear implant.
The cochlear implant is a transformative technology that allows children to be mainstreamed into regular schools, adults to have a wide range of job opportunities, and all recipients to connect in new and important ways with their families, friends, and society at large. The resulting human and economic benefits are immense.
In many parts of the world, cost is a barrier to widespread applications of the technology, even though the benefits ultimately far outweigh the cost. The principal expenses are in providing the appropriate medical infrastructure and care. The cost of the device also plays a role, but that cost is coming down and is not the dominant factor for most countries. Several of us in this room are working to reduce or remove the cost barrier, and to improve hearing health care worldwide, which includes prevention, screening, and treatments in addition to cochlear implants.
This magnificent award will greatly increase awareness of how cochlear implants can enable severely and profoundly deaf persons to realize their full potential in life, and that awareness will in turn facilitate further dissemination and development of this marvelous technology. Thank you for welcoming Graeme, Ingeborg, and me into the Lasker family, and thank you for the highly favorable tailwind you have given us and our colleagues to do more!
Interview with Richard H. Scheller and Thomas C. Südhof
Video Credit: Susan Hadary
