
Victor R. Ambros
University of Massachusetts Medical School

David C. Baulcombe
University of Cambridge

Gary B. Ruvkun
Harvard Medical School, Massachusetts General Hospital
For discoveries that revealed an unanticipated world of tiny RNAs that regulate gene function in plants and animals.
The 2008 Albert Lasker Award for Basic Medical Research honors three scientists who discovered an unanticipated world of tiny RNAs that regulate gene activity in plants and animals. Victor R. Ambros (University of Massachusetts Medical School, Worcester) and Gary B. Ruvkun (Massachusetts General Hospital, Boston, Harvard Medical School) unearthed the first example of this type of molecule in animals and demonstrated how the RNAs turn off genes whose activities are crucial for development. David C. Baulcombe (University of Cambridge) established that small RNAs silence genes in plants as well, thus catalyzing discoveries of many such RNAs in a wide range of living things. His findings led to the identification of the biochemical machinery that unifies numerous processes by which small RNAs govern gene activity.
Ambros, Baulcombe, and Ruvkun did not set out to unveil small regulatory RNAs. Ambros and Ruvkun were studying how the worm Caenorhabditis elegans develops from a newly hatched larva into an adult. Baulcombe, in a seemingly unrelated line of inquiry, was probing how plants defend themselves against viruses. All three investigators possessed the open mindedness, wisdom, and experimental finesse to entertain the possibility — and then verify — that tiny RNAs could perform momentous feats. Their work has led to the realization that these molecules are pivotal regulators of normal physiology as well as disease.
RNA — the little molecule that could
In the early 1980s, Ambros joined the laboratory of Robert Horvitz at the Massachusetts Institute of Technology as a postdoctoral fellow. He wanted to outline the means by which genes choreograph the construction of fully formed adults from single cells. Analyses of flies had revealed that certain genes instruct embryos where to place body parts — for example, wings belong on each side and legs belong on the bottom. But Ambros was intrigued by the notion that other genes might specify the timing — rather than the location — of developmental events; alterations in such genes might cause cells and tissues to adopt fates that are normally associated with earlier or later stages of development.
He directed his attention toward one of the first-known genes of this type, called lin-4, which had been identified earlier in the laboratory of Sydney Brenner (Lasker Special Achievement Award, 2000) and subsequently characterized by Horvitz, Martin Chalfie, and John Sulston. Ambros recognized that, during worms’ trek toward adulthood, those with inactive lin-4 get stuck repeating early larval stages. Consequently, they lack cell types and structures typical of fully formed animals and instead contain extra copies of cells ordinarily produced only at early stages. These observations suggested that normal lin-4 allows immature worms to advance past a particular developmental stage; animals with the defective version cannot overcome that hurdle. Ambros discovered that worms lacking a different gene — lin-14 — were the antithesis of those with inactive lin-4. The animals skip early steps in development and prematurely acquire characteristics that normally appear later. These and other results suggested that lin-4 and lin-14 exert opposite effects in worm cells.
To dig further into lin-14’s function and its possible relationship with lin-4, Ruvkun, who by this time (1982) had joined Horvitz’s laboratory as a postdoctoral fellow, collaborated with Ambros to isolate the lin-14 gene. After the investigators set up independent laboratories in the mid 1980s, Ruvkun, at Massachusetts General Hospital in Boston, established that the protein product of lin-14 is abundant during early larval stages and then its quantities plummet. Under conditions in which it unnaturally remains plentiful, early steps repeat, suggesting that the normal drop in the lin-14 protein allows worms to proceed to later stages. Ambros, at Harvard University, found that lin-4 dampens lin-14 activity and thus a picture emerged about how the genes collaborate. At the appropriate time, lin-4 blocks lin-14 and thus allows worms to continue their developmental trajectory.
Ruvkun sought to identify the portion(s) of lin-14 that lin-4 targets, so he tracked down certain genetic anomalies in lin-14’s sequence that underlie excess production of the lin-14 protein. He found that these alterations reside in the area of the gene that follows the protein blueprint, a span called the 3′ untranslated region (3′ UTR). The perturbations do not influence amounts of the protein’s messenger RNA (mRNA), the molecule that carries genetic information from DNA to the cell’s protein-making factory, Ruvkun showed. Rather, they alter protein quantities. Therefore, molecules that turn off lin-14 after early stages of development presumably exert their effects through the 3′ UTR region of the lin-14 mRNA and prevent the cell from translating its code into protein.
In the meantime, Ambros’s laboratory was isolating the lin-4 gene, which they assumed encoded a protein; although a few RNAs were known to control gene activity in bacteria, conventional wisdom held that, in animal cells, proteins alone enjoy such powers. The team homed in on smaller and smaller pieces of DNA from normal animals that restore typical developmental behavior to a worm that lacks lin-4. Stretches of DNA that were far shorter than standard genes worked. Eventually, the researchers began considering the possibility that its product was an RNA, but they still assumed that the regulatory molecule they sought would be a respectable size. The smallest RNAs known to do anything important in cells contained about 75 nucleotide (nt) building blocks. Eventually, however, their experiments led them to a tiny RNA, composed of about 22 nucleotides. A larger — 61 nt — molecule that contained the smaller RNA appeared as well and Ambros noticed that it could fold into a double-stranded ‘hairpin’ — a structure whose significance would become clear years later.
In an exciting exchange of data, Ambros and Ruvkun realized that the 22-nt lin-4 RNA matched sections within the 3′ UTR of the lin-14 mRNA: These sequences could bind one another by the same base-pairing rules that hold together the Watson and Crick DNA strands. In this view, the tiny lin-4 RNA settles on the target lin-14 mRNA — in its 3′ UTR — and the resulting double-stranded structure somehow interferes with translation of the lin-14 mRNA’s genetic information into protein (see illustration).
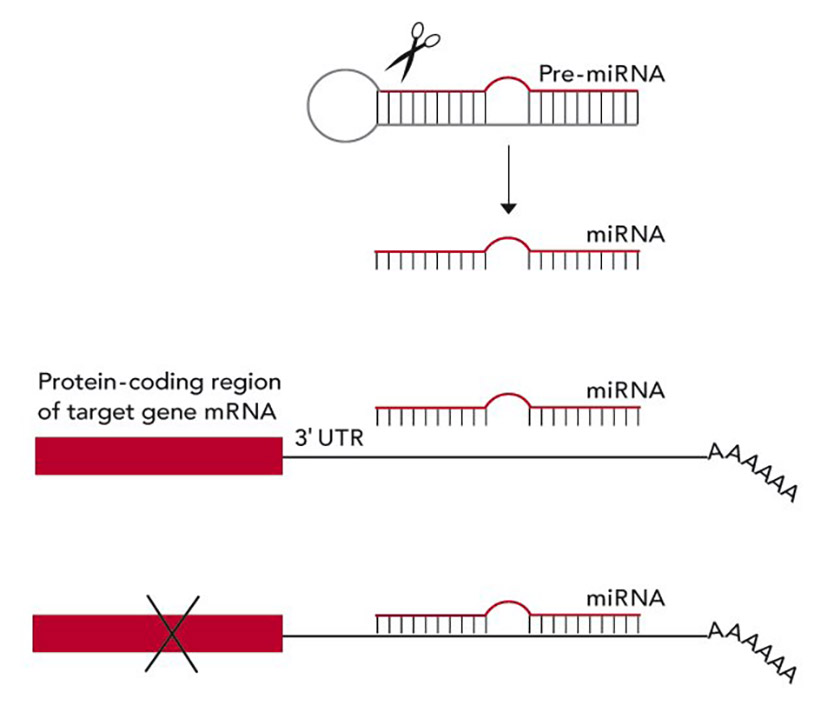
Small but mighty. This scheme shows how one type of tiny RNA, a microRNA (miRNA), silences genes. It is cut out of a precursor hairpin-shaped pre-miRNA to form a mature miRNA, which binds to the 3′ untranslated region (3′ UTR) of a target gene’s messenger RNA and turns off its activity. [Credit: Carin Cain. Based on an illustration from Victor Ambros]
Branching out to plants and beyond
Across the Atlantic, David Baulcombe, then of the Sainsbury Laboratory in Norwich, UK, was studying how plants resist viruses. When he and others added to viral-infected plants unusual versions of viral genes, the mRNA copies of the normal genes as well as the newly introduced ones disappeared. Similarly, experimentally added non-viral genes suppressed activity of plant genes that contained similar sequences. Baulcombe proposed that such gene silencing occurs when RNAs embrace target mRNA — through typical Watson-Crick base-pairing — and promote destruction of the mRNA or interfere with its translation into protein. However, no one could find such RNAs.
Baulcombe reasoned that the predicted RNAs might have eluded researchers because the molecules were shorter than anyone imagined and thus, experiments had not been designed to detect them. In 1999, he and a postdoctoral fellow in his laboratory, Andrew Hamilton, devised a hunt specifically for small RNAs. They added test genes to plants and found 25-nt-long RNAs that matched; furthermore, these small RNAs appeared only under conditions in which target mRNA activity was shut off. The stunning similarity in size between the plant and worm RNAs suggested that small regulatory RNAs exist in many organisms. Furthermore, it hinted at the presence of cellular machinery that dedicates itself to creating these precisely sized molecules and then uses them to quash gene activity.
In 2000, Ruvkun’s laboratory discovered a second tiny regulatory RNA in worms of exactly the same size as the lin-4 RNA and in the same genetic pathway. Similar to the lin-4 RNA, this let-7 RNA dampens activity of its target gene through its 3′ UTR. Furthermore, its sequence too resides within a larger molecule that folds up on itself to form a double-stranded hairpin structure. Later that year, Ruvkun found that many other creatures, including humans, fruit flies, chickens, frogs, zebrafish, mollusks, and sea urchins, carry their own versions of let-7, which could also fold into hairpins. The apparent binding site for let-7 RNA in its target was conserved in some of these organisms as well. Moreover, let-7 RNA appeared and disappeared at similar points during development in many of the animals.
The small RNAs, now called microRNAs (miRNAs), had broken through their designation as “worm curiosities.” Researchers realized that the miRNAs likely execute vital functions during growth and development of other creatures as well. Multiple teams raced to expose regulatory RNAs of approximately 22 nucleotides in length. In 2001, Ambros’s group, now at Dartmouth Medical School, in Hanover, as well as those of David Bartel (Massachusetts Institute of Technology) and Thomas Tuschl (Max Planck Institute for Biophysical Chemistry, Göttingen) discovered almost 100 of these small regulatory RNAs in flies, humans, and worms.
In addition to revealing that small regulatory RNAs dwell in organisms other than worms, Baulcombe’s finding caught many researchers’ attention because it seemed related to a process called RNA interference (RNAi), which had recently exploded onto the biological scene. In RNAi, long RNAs injected into cells hamper gene activity from similar sequences. No one knew why organisms possessed this ability, but presumably it played some role in normal physiology. In 1998, Andrew Fire (Carnegie Institution of Washington, Baltimore) and Craig Mello (University of Massachusetts Medical School, Worcester), published a watershed paper that defined the fundamental features of RNAi (which garnered them the Nobel Prize in 2006). That work yielded the surprising insight that the process depends on double-stranded RNA. However, the means by which double-stranded RNA triggered silencing remained mysterious.
Experiments from Baulcombe’s laboratory provided the crucial clues. Production of the silencing RNA strand depended on the presence of the other strand, he had noticed. This observation suggested that, at some point during manufacture of the small regulatory RNA, it exists as part of a double-stranded molecule. Suddenly it seemed possible that Baulcombe’s tiny RNAs arose by trimming longer molecules of the type that Fire and Mello had discovered. Furthermore, this notion suggested that the hairpin-like lin-4 and let-7 RNAs similarly gave rise to the mature, 22-nt entities.
Scientists wondered whether the cell deployed the same biochemical machinery to create and use RNA molecules that subdued gene activity in all of these gene-silencing systems. However, the mechanisms of the worm miRNAs seemed to differ from those of the plant molecules as well as RNAi. Unlike the system that Ambros and Ruvkun had been untangling, which allowed mRNA to accumulate but thwarted cells’ abilities to translate the information it contained into protein, the plant system and RNAi destroyed mRNA. For that reason and others, many people doubted that the processes were connected. Still the possibility that they shared a common mechanism and machinery tantalized researchers.
In 2001, the Mello, Ruvkun, and Fire groups collaborated to show that efficient liberation of the lin-4 and let-7 RNAs from the hairpin molecules relies on the C. elegans version of Dicer, an enzyme that Gregory Hannon (Cold Spring Harbor Laboratory) discovered and named for its ability to chop dsRNA into uniformly sized, small RNAs that direct mRNA destruction during RNAi. These results and others, including similar ones generated by Philip Zamore (University of Massachusetts Medical School, Worcester), cemented the connection between miRNAs and RNAi, thus providing one biological ‘reason’ for the RNAi machinery. Moreover, they identified the apparatus by which cells generate miRNAs and harness them for key pursuits.
Studies in the past several years have indicated that the human genome contains more than 500 and perhaps as many as 1000 miRNAs that could collectively control a third of all of our protein-producing genes. These regulatory molecules have been implicated in a wide range of normal and pathological activities. They play roles not only in embryonic development, but in blood-cell specialization, cancer, muscle function, heart disease, viral infections, and possibly neurological signaling and stem-cell behavior. Researchers are exploring the possibility of using miRNAs ‘signatures’ for diagnosis and prognosis and are considering manipulating their quantities for therapeutic purposes.
Looking where no one had looked before, Ambros, Baulcombe, and Ruvkun spied an unforeseen universe of potent molecules. Their work has elevated these hitherto unrecognized agents into the spotlight of biology and medicine.
by Evelyn Strauss
Key publications of Victor Ambros
Ambros, V. (1989). A hierarchy of regulatory genes controls a larva-to-adult developmental switch in C. elegans. Cell. 57, 49-57.
Lee, R., Feinbaum, R., and Ambros, V. (1993). The heterochronic gene lin-4 of C. elegans encodes small RNAs with antisense complementarity to lin-14. Cell. 75, 843-854.
Moss, E., Lee, R., and Ambros, V. (1997). Control of developmental timing by the cold shock domain protein Lin-28 and its regulation by the lin-4 RNA. Cell. 88, 637-646.
Olsen, P.H., and Ambros, V. (1999). The lin-4 regulatory RNA controls developmental timing in C. elegans by blocking LIN-14 protein synthesis after the initiation of translation. Dev. Biol. 216, 671-680.
Lee, R.C. and Ambros, V. (2001). An extensive class of small RNAs in Caenorhabditis elegans. Science. 294, 862-864.
Lee, R., Feinbaum, R., and Ambros, V. (2004). A short history of a short RNA. Cell. S116, S89-S92.
Key publications of David Baulcombe
Brigneti, G., Voinnet, O., Li, W.X., Ji, L.H., Ding, S.W., and Baulcombe, D.C. (1998). Viral pathogenicity determinants are suppressors of transgene silencing in Nicotiana benthamiana. EMBO J. 17, 6739-6746.
Dalmay, T., Hamilton, A.J., Rudd, S., Angell, S., and Baulcombe, D.C. (2000). An RNA-dependent RNA polymerase gene in Arabidopsis is required for posttranscriptional gene silencing mediated by a transgene but not by a virus. Cell 101, 543-553.
Hamilton, A.J. and Baulcombe, D.C. (1999). A species of small antisense RNA in posttranscriptional gene silencing in plants. Science. 286, 950-952.
Hamilton, A.J., Voinnet, O., Chappell, L., and Baulcombe, D.C. (2002). Two classes of short interfering RNA in RNA silencing. EMBO J. 21, 4671-4679.
Ratcliff, F., Harrison, B.D., and Baulcombe, D.C. (1997). A similarity between viral defense and gene silencing in plants. Science. 276, 1558-1560.
Baulcombe, D.C. (2006). Short silencing RNA: The dark matter of genetics? Cold Spring Harb. Symp. Quant. Biol. 71, 13-20.
Key publications of Gary Ruvkun
Ruvkun, G. and Giusto, J. (1989). The Caenorhabditis elegans heterochronic gene lin-14 encodes a nuclear protein that forms a temporal switch during development. Nature. 338, 313-319.
Wightman, B., Bürglin, T.R., Gatto, J., Arasu, P., and Ruvkun, G. (1991). Negative regulatory sequences in the lin-14 3′ untranslated region are necessary to generate a temporal switch during C. elegans development. Genes Dev. 5, 1813-1824.
Wightman, B., Ha, I., and Ruvkun, G. (1993). Post-transcriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell. 75, 855-862.
Reinhart, B.J., Slack, F.A., Basson, M., Pasquinelli, A.B., Bettinger, J.C., Rougvie, A.C., Horvitz, H.R., and Ruvkun, G. (2000). The 21 nucleotide let-7 RNA regulates C. elegans developmental timing. Nature. 403, 901-906.
Pasquinelli, A., Reinhart, B., Slack, F., Maller, B., Kurodo, M., Martindale, M., Srinivasan, A., Fishman, M., Hayward, D., Ball, E., Degnan, B., M�ller, P., Spring, J., Finnerty, J., Corbo, J., Levine, M., Leahy, P., Davidson, E., and Ruvkun, G., (2000). Conservation across animal phlylogeny of the sequence and temporal expression of the 21 nucleotide let-7 heterochronic regulatory RNA. Nature. 408, 86-89.
Ruvkun, G., Wightman, B., and Ha, I. (2004). The 20 years it took to recognize the importance of tiny RNAs. Cell. S116, S93-S96.
Award presentation by H. Robert Horvitz
The cell is the fundamental unit of life. Our bodies are made up of cells, about 10 trillion cells, and these cells are of many types: skin cells, blood cells, muscle cells, nerve cells, and so on. These cell types are strikingly different from each other, yet the genes they carry are, in almost all cases, identical. How can this be? The answer is that the expression of genes is controlled, so that some genes are expressed in some cells and other genes are expressed in other cells. The consequent difference in gene expression is what makes cells differ. Abnormalities in gene expression are responsible for many diseases, for example, many cancers. How gene expression is controlled is one of the fundamental and most important problems in biology.
Acceptance remarks

Acceptance remarks, 2008 Lasker Awards Ceremony
On my application for admission to MIT, my essay on the topic of “Why I want to attend MIT,” consisted of just six words: “I want to be a scientist.” My path, from that state of extreme naiveté in 1970 to this occasion today, would be impossible without an enormous web of support, consisting of my family, friends, teachers, mentors, bosses, colleagues, collaborators, and my wife Candy Lee. When I applied to MIT, I knew that if I hoped to become a scientist, I needed to be in the right place, but I did not know then that it would matter so much that I should be among the right people. This award is really in recognition of a confluence of events and people in my life, and it is my enormous pleasure that so many of those who have helped me are here today. In the spirit of enforced brevity, I will just say that you know who you are, and I refer everyone to my written comments for more details (and I should also mention a newcomer to my support net, Mike Czech, who hired me into my latest and best job, and who has become my friend and mentor).
I think my six-word MIT admissions essay was an attempt to distill a poorly defined adolescent dream — of belonging one day to a grand tradition populated by mythic figures: Einstein, Hubble, Galileo. But, what I learned at MIT, to my relief and delight, was that science is actually done by regular folks! To this day, what I love most about science is that it is such a deeply, intensely human enterprise. The success of the enterprise of science, and of the individual scientist, is derived precisely from the fact that we do it together. We work together in synergy as small teams, such as Candy Lee and Rhonda Feinbaum did to discover the first microRNA; we communicate our treasured, secret data across lab borders, as Gary Ruvkun and I did to enable us to discover the antisense base-pairing between microRNAs and targets; and we publish to the world our findings, so that for example, I opened a journal in 1999, and was astonished by a report from David Baulcombe’s group of little RNAs, just like lin-4, way over there in plants.
Not only do we do science as individuals, teams, and as a community of scientists, perhaps most importantly, science is an enterprise of and by the public. One does not have to be a scientist to contribute substantively to scientific discovery. We can be certain that without financial support from foundations and public agencies, particularly the National Institutes of Health, and without the commitment of universities to basic research, we would not be here today, and nobody would know anything about small RNAs, or about so many other things. For me, the Lasker Award is a profound personal honor, because I know that it recognizes not just the creative effort of Candy and Rhonda, Gary and his group, David and his co-workers, but it also emphasizes once again the vital legacy of Mary Lasker and her pioneering work on behalf of publicly supported science in our modern era. Science is among the best things that we do as a species, and I am proud and happy beyond words to be part of it.

Acceptance remarks, 2008 Lasker Awards Ceremony
My initial reactions to hearing about the Lasker award were surprise, delight, satisfaction, gratitude to colleagues, and a nice warm feeling. I was also pleased that there had been recognition of plants as model systems — I have written about that in the commentary article. After a while, I also started wondering how I got into this prize-winning position. Part of the answer of course is luck and having great colleagues. The other part is because EVENTUALLY we — my group — did what we should do as scientists. Through a process of deduction and controlled experimentation we worked out that the important molecules are antisense of the target molecule and they are small and finally we made some progress.
Now the point I would like to make is about the word “EVENTUALLY” — the whole process took some considerable time, and it was a while before we even realized that we were looking at something extraordinary. Even then, we messed about with misguided models based on proteins or even DNA interactions before we got on the right track. What we did initially was what all people do most of the time — we were jumping to conclusions. The right side of our brain took over and our thinking was based on preconceptions, hunches, and incomplete information.
In everyday life, the intuitive right hemisphere process serves us quite well: it would be a pretty strange and feeble existence without it. Right hemisphere thinking may even have a proper place in the early pathway of a scientific discovery. But, for most of the time in science, the evidence-based logical left hemisphere should dominate.
Now, the point of all of this is that practically everyone on the planet has a functioning left hemisphere, and potentially everyone could embrace the scientific way of thinking from time to time, but they do not. Perhaps if people could understand that many scientists struggle with this right and left hemisphere conflict, it would be easier for them to accept science and what it has to offer. I see getting widespread acceptance of science and its technological opportunities as being a significant challenge — even now. The Lasker Foundation has established an excellent forum to promote science and I am honored to be associated with it.

Acceptance remarks, 2008 Lasker Awards Ceremony
The discovery of a previously unsuspected civilization of tiny RNAs was the first perfect storm of the genome era. I would like to acknowledge the inspired discoveries of my co-awardees Victor Ambros and David Baulcombe, whom I have now known for most my scientific career. It is important for those interested in medical research to realize that much of the tiny RNA revolution emerged from non-mammalian genetic analysis — from plant biology, from worm genetics, from the fungi, from the protozoa. Much of this research was funded generously over the past decade or two, reflecting well on the wisdom of the many peer review systems, private and public, which support science. It is important to continue to explore the diversity of biology; our next revolutions, scores of them, are likely to emerge from here.
It was Sputnik in 1957 that launched me into science. The ensuing US launches were fully televised and breathlessly reported. Television was a new medium, and unlike today, with the inevitable launch delays, there could be four or five hours of continuous network coverage, with hours of prerecorded interviews with bowtied, flattop haircut scientists to fill the time. I fondly remember the excitement of Walter Cronkite and Jules Bergman when they interviewed the engineers who designed space capsules or the scientists who described orbital mechanics. The exciting concepts that I learned from television those few dozen mornings from 1961 to 1966 were far more sophisticated than what was being taught in my elementary school and were key elements in shaping me.
I thank my parents for my first microscope at age 5, for my telescope at age 8, for taking me to the Oakland Public Library once a week to check out a dozen books, for teaching me how a steel mill works, for my first ham radio, for their celebration of Berkeley as Athens, for the awe in their voices when they said, “nuclear physicist.”
One of the most wonderful dividends of this sort of recognition is that I can interpret aspects of my autobiography, exploring how my past might have helped me come to discoveries. And that there are audiences that I can try to interest in my saga. It is at least possible that being a Jew in Oakland, California, made me enough of an outsider that I did not neatly fit into any community and escaped a sense of entitlement. And that my kinship with the great Jewish comics of the ’50s and ’60s fostered a certain irreverance and a warped perspective,and made me funny so that people might gravitate to work with me. It is possible that my undergraduate training in physics taught me not quantitative reasoning but rather how a scientific revolution at its very beginning can be recognized. And that my year working in a tree-planting cooperative in Oregon after graduating from college taught me the joys of a crew working together, cooperatively, on a hard problem. And that my year of travel on third-class buses from Mexico to Tierra del Fuego, with my encounters with thousands of strangers, may have trained me in the instantaneous assessment of the others, no doubt helping me to recruit the most interesting and intelligent students and post-docs to my lab.
I arrived at graduate school in 1976, with no idea how science is done. From my fellow graduate students and post-docs at Harvard and MIT, I learned how to eat and breathe science, how to be thrilled with discoveries at the moment of their presentation. From my teachers, Fred Ausubel, Wally Gilbert, and Bob Horvitz, all of whom are here today, I learned something more intangible but key: good taste in picking scientific problems, and by their examples, how to unravel the tangled webs that life has evolved.
So the tribes of my education, my childhood, my lab, and my academic environment were uniquely inspiring, supportive, and loads of fun. But my home tribe has been the wellspring of strength and joy. And I am truly grateful to Natasha Staller, and our daughter Victoria, for the joyous home life of our little tribe.
Interview with Victor R. Ambros, David C. Baulcombe, and Gary B. Ruvkun
Video Credit: Susan Hadary
