
Karl Deisseroth
Stanford University

Peter Hegemann
Humboldt University of Berlin

Dieter Oesterhelt
Max Planck Institute of Biochemistry
For the discovery of light-sensitive microbial proteins that can activate or silence individual brain cells and for their use in developing optogenetics—a revolutionary technique for neuroscience
The 2021 Albert Lasker Basic Medical Research Award honors three scientists for the discovery of light-sensitive microbial proteins that can activate or silence individual brain cells and for their use in developing optogenetics, a revolutionary technique for neuroscience. Dieter Oesterhelt (Emeritus, Max Planck Institute of Biochemistry, Martinsried) uncovered an archaeal protein that pumps protons out of cells when illuminated. Peter Hegemann (Humboldt University of Berlin) then found related channel proteins in single-celled algae. Karl Deisseroth (Stanford University) harnessed these molecules to create light-triggered systems that can be deployed in live, free-moving animals to decipher the role of specific classes of—and even individual—neurons within labyrinthine brain circuits. Hundreds of laboratories around the world are now using this approach, called optogenetics, to untangle the elaborate networks that underlie healthy physiology and neurological disease.
Each of our 86 billion neurons forms, on average, a thousand connections with other neurons. Electrical and chemical signals among them produce the intricate message patterns that drive sensations, thoughts, feelings, and behaviors. To elucidate how the brain runs smoothly and how it sputters to foment illness, researchers need to unravel a mind-boggling wiring puzzle.
For decades, scientists have fantasized about arousing or quelling select neurons to assess the consequences and thereby discern the cells’ normal functions. Traditional methods fall short because they act too slowly or affect all neurons in large swathes of the brain. An ideal strategy would facilitate rapid manipulation of brain circuits in alert animals with locational and cell-type precision. In 1999, Francis Crick speculated that light might lend itself to this venture, although he deemed the idea “rather far fetched.”
Pumped up by a purple membrane
In 1969, Oesterhelt went on sabbatical from the University of Munich to the late Walter Stoeckenius’s lab (University of California, San Francisco). Stoeckenius was studying the membranes that surround a microbe now known as the archaeon Halobacterium salinurum, which flourishes in high salt environments. This creature falls apart in low salt solutions to generate separable biochemical portions, including a purple membrane fraction.
Adding particular solvents made the color fade, Oesterhelt noticed. He was perplexed by this chemically confounding behavior, but when a colleague told him that frog eyes bleach from red to yellow upon overillumination, he had a preposterous and exhilarating thought. Maybe the purple material in the single-celled archaeon contained retinal, the vitamin A-based compound that joins with proteins called opsins. Retinal and opsins are essential for vision in animals.
Oesterhelt established that the sole protein in the purple membrane binds retinal, and he and Stoeckenius named it bacteriorhodopsin after its well-known vertebrate cousin rhodopsin. In 1971, they proposed that bacteriorhodopsin functions as a light sensor, or photoreceptor.
Oesterhelt, back in Germany, and Stoeckenius continued to work in parallel and as collaborators. Oesterhelt wondered if a chemical change accompanied the color shift, so he exposed membrane preparations and intact cells to light and measured the pH. It plummeted, indicating that hydrogen ions, or protons, escaped into solution. This experiment and others established that bacteriorhodopsin transports protons out of the cell.
Subsequent studies by Oesterhelt and others established that retinal plays a pivotal role. When light hits retinal, it absorbs a photon and snaps into a different shape that forces the bacteriorhodopsin protein’s innards to change their relative positions. This perturbation places a particular proton into a chemically unfriendly environment. Consequently, it jumps to a more welcoming spot, and eventually a proton winds up outside the cell. Analogous events replenish the lost proton with one from the cell’s interior, restoring bacteriorhodopsin’s original configuration. In this way, a photon instigates transfer of a single proton from inside to outside the cell.
Scientists later found a second type of retinal-containing light-activated pump in Halobacteria. This one, called halorhodopsin, conveys chloride ions into cells. Oesterhelt’s landmark biophysical, structural, and genetic studies of bacteriorhodopsin and halorhodopsin provided foundational insights that informed the eventual development of optogenetics.
Channeling ion flow: a new class of photoreceptors
In 1980, Hegemann joined Oesterhelt (by then at the Max Planck Institute for Biochemistry, Martinsried) as a PhD student. Soon afterward, some papers about the single-celled alga Chlamydomonas caught Hegemann’s eye. This organism—a eukaryote, like mammals—has an eyespot that enables it to swim toward conditions that optimally support photosynthesis, and new evidence hinted that the eyespot’s light-sensing instrument relies on retinal.
Intrigued, Hegemann decided to track down the photoreceptor’s presumptive protein component when he established his own lab at the Max Planck Institute of Biochemistry, Martinsried. In 1991, he found that ion currents into the organism rise extremely rapidly in response to light, much faster than analogous events in the visual pathway of higher animals, whose rhodopsins require other molecules to exert their effects. Bolstered by additional results several years later, he reasoned that insufficient time existed for a molecular signal to pass from the light detector to the ion channel and proposed that the two elements form a single protein.
To test his all-in-one hypothesis, he needed a way to interrogate the apparatus’s properties in a clean system. In 2000, other scientists compiled a collection of DNA sequences from Chlamydomonas. Hegemann (by this time at the University of Regensburg) identified one that encodes a protein that resembles bacteriorhodopsin—most notably at the spots that interact with retinal and compose its proton-transporting network.
Hegemann, in collaboration with Georg Nagel and Ernst Bamberg (Max Planck Institute for Biophysics, Frankfurt), put the gene into frog eggs and evaluated electrical behavior produced by the protein encoded by the introduced gene. As Hegemann had hypothesized, this protein acts as a channel that opens upon illumination and allows protons to rush in. They announced these discoveries in 2002. The following year, they reported similar data for a second light-activated algal ion channel, which admits in addition positive ions such as sodium and calcium. Furthermore, they demonstrated that it can function when placed in mammalian kidney cells. They dubbed this new class of proteins channelrhodopsins (ChRs) (Figure 1).
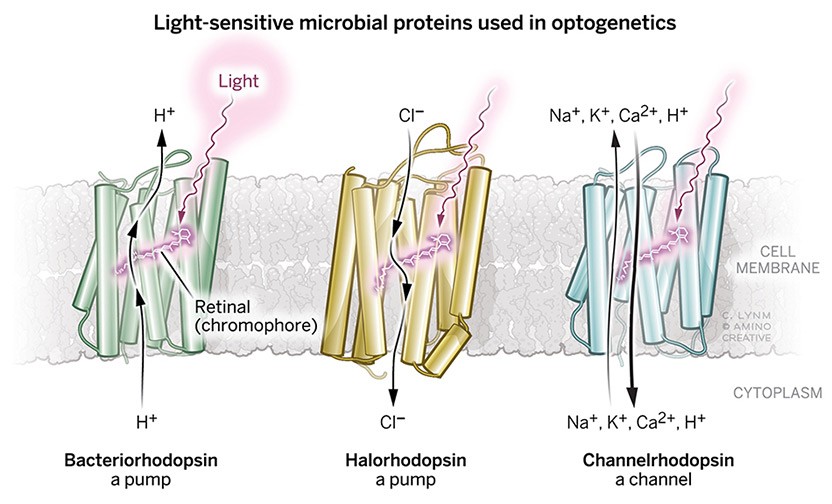
Micro origins, macro impact
These three proteins, whose discovery and utility gave rise to optogenetics, facilitate light-triggered ion transfer across cell membranes. The archaeon Halobacterium salinarum contains bacteriorhodopsin, which pumps protons out of cells, and halorhodopsin, which pumps chloride ions into cells. The single-celled algae Chlamydomonas reinhardtii contains channelrhodopsins; when these channels open, positive ions surge through the membrane.
Illustration: Cassio Lynm/© Amino Creative
A bright idea
These findings electrified neurobiologists, including Deisseroth. An influx of positive ions is exactly what sparks neuronal signals, and the prospect of using a single molecule to direct brain circuits was captivating. By this time, others, including Gero Miesenböck (University of Oxford), had attempted to control neurons with light, but those procedures were impractical because, for example, they required simultaneous delivery of multiple components.
Deisseroth and Stanford colleagues Edward Boyden and Feng Zhang, in collaboration with Nagel and Bamberg, engineered a harmless virus to deliver a ChR gene to neurons grown in culture dishes. Light exposure provoked ion flow that was fast, adjustable, reproducible—and strong enough to prompt neuronal firing. They published these observations in 2005, and over the following year, four other groups described similar results, including scientists from Case Western Reserve University, Humboldt University, Frankfurt’s Goethe University, Sendai’s Tohoku University, and Wayne State University. Soon afterward, many labs across the globe began using the technology.
To implement the scheme in the mammalian brain, though, multiple innovations were required, and Deisseroth spearheaded these advances. Researchers had to devise a way to deliver light that would penetrate tissue safely, for example, and no one knew whether mature mammalian brains contain the essential retinal cofactor.
In 2007, Deisseroth reported not only that the technology operated in live rodents, but also that it could command behavior (Figure 2). He delivered a ChR gene to a part of the brain that governs whisker movement. When he shot blue light through a thin optical fiber that he had implanted, their whiskers twitched. Because the rodents’ heads were held in place, the experimental setup was not of general utility, but later that year, Deisseroth showed that the same approach worked on mice that could move freely.
Many improvements followed, as researchers honed known microbial opsins for various purposes and discovered others. In 2008, Deisseroth and Hegemann collaborated to find a natural ChR that responds to yellow rather than blue light. Later, they designed ChRs that spur especially strong currents and others that remain on for long periods of time and can be switched off with a different color of light.
In 2009, Deisseroth’s student Viviana Gradinaru pioneered another impactful refinement. Because each neuron extends its tentacle-like axons throughout the brain, the projections reach disparate locations. By focusing light in a particular area, the investigators selectively controlled firing by opsin-containing neurons whose cell bodies might lie far away. The team used this method on mice with a Parkinson Disease-like condition to probe how deep brain stimulation delivers its benefits (Alim Louis Benabid and Mahlon R. DeLong, Lasker Clinical Research Award, 2014).
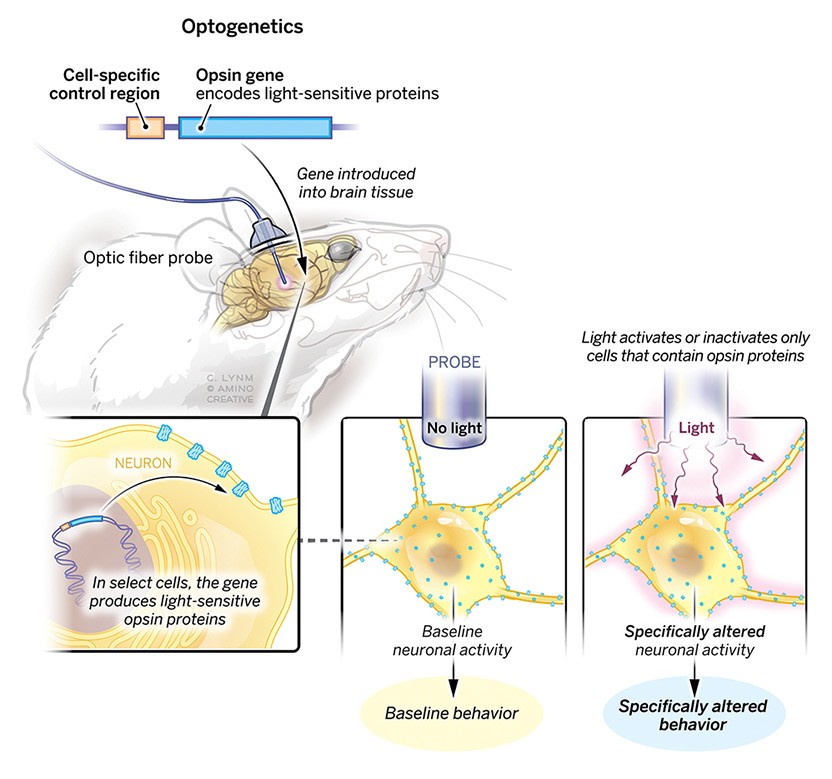
Flashes for insight
In the standard optogenetic strategy, scientists engineer a control region that turns on the adjacent opsin gene in certain cell types. This DNA is packaged into a benign virus, which delivers it to the brain, where opsin protein is produced only in the selected neurons. An optic fiber carries light, which activates the opsins, thus turning these specific neurons on or off and allowing researchers to assess processes of interest.
Illustration: Cassio Lynm/© Amino Creative
Enlightening technology
For many years, the only channels available to the optogenetics field turned neurons on, as they allowed positive ions such as sodium to stream in. Some of the opsin pumps transported negative ions into cells, but because only one ion moved at a time, neuronal inhibition was weak.
In 2012, Osamu Nureki (University of Tokyo) and Deisseroth, in collaboration with Hegemann, presented the first high-resolution structure of a ChR, and they saw that the ionic travel pathway is lined with amino acids that repel negative ions. Guided by this information, Deisseroth and Hegemann independently re-engineered the channel so it would transport the negative chloride ion. This new tool blocked neuronal firing; thus, it dramatically amplified the experimental power of optogenetics by enabling inhibition with channels.
Turning on and off genetically defined subsets of neurons in living animals has transformed the study of brain function, but cells of a given type don’t always act together. In 2012, Deisseroth’s team managed to control single neurons in live mice by customizing opsins for use with so-called two-photon technology. Seven years later, the investigators applied this system to manipulate single cells in an area of the mouse brain associated with rewards gained from social interactions and food consumption. For example, they restrained feeding by targeting certain social cells.
Scientists are applying optogenetics to tease apart a vast number of adaptive and maladaptive behaviors (Figure 3). By using inhibitory and stimulatory opsins, Deisseroth and colleagues stopped thirsty mice from seeking water and prodded fully hydrated animals to guzzle. Similarly, investigators are homing in on the mechanism by which leptin regulates body weight (Douglas Coleman and Jeffrey M. Friedman, Lasker Basic Research Award, 2010), identifying some neurons that compel animals to eat voraciously and others that suppress appetite.
Complexity decoded
Optogenetic has already yielded rich information about numerous behavioral and physiological phenomena in experimental animals. It is enabling scientists to determine the contribution of individual neurons to these activities.
Illustration: Cassio Lynm/© Amino Creative
The strategy can also be applied to more complex cognitive phenomena. Deisseroth optogenetically dissected anxiety into its physiological, behavioral, and emotional components, and other researchers have similarly broken down mouse parenting into a circuit that propels a mother to find her babies and another that drives her to care for them.
The synergistic discoveries of Oesterhelt, Hegemann, and Deisseroth have launched the Era of Optogenetics—an age in which re-engineered retinal-containing opsins from single-celled organisms allow researchers to peer into the brains of living animals. Scientists can now study neurons and their circuitry with stunning clarity and probe their functional underpinnings with unprecedented resolution. Optogenetics has already exposed hitherto unknown secrets and it promises to reveal additional elusive ones.
by Evelyn Strauss
Selected Publications–Discovery of Microbial Opsins
Oesterhelt, D., and Stoeckenius, W. (1971). Rhodopsin-like protein from the purple membrane of Halobacterium halobium. Nat. New Biol. 233, 149-152.
Oesterhelt, D., and Stoeckenius, W. (1973). Functions of a new photoreceptor membrane. Proc. Natl. Acad. Sci. USA. 70, 2853-2857.
Bamberg, E., Tittor, J., and Oesterhelt, D. (1993). Light-driven proton or chloride pumping by halorhodopsin. Proc. Natl. Acad. Sci. USA. 90, 639-643.
Kolbe, M., Besir, H., Essen, L.-O., and Oesterhelt, D. (2000). Structure of the light-driven chloride pump halorhodopsin at 1.8 Å resolution. Science. 288, 1390-1396.
Nagel, G., Ollig, D., Fuhrmann, M., Kateriya, S., Musti, A.M., Bamberg, E., and Hegemann, P. (2002). Channelrhodopsin-1, a light-gated proton channel in green algae. Science. 296, 2395-2398.
Nagel, G., Szellas, T., Huhn, W., Kateriya, S., Adeishvili, N., Berthold, P., Ollig, D., Hegemann, P., and Bamberg, E. (2003). Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proc. Natl. Acad. Sci. USA. 100, 13940-13945.
Kato, H.E., Zhang, F., Yizhar, O., Ramakrishnan, C., Nishizawa, T., Hirata, K., Ito, J., Aita, Y., Tsukazaki, T., Hayashi, S., Hegemann, P., Maturana, A.D., Ishitani, R., Deisseroth, K., and Nureki, O. (2012). Crystal structure of the channelrhodopsin light-gated cation channel. Nature. 482, 369-374.
Wietek, J., Wiegert, J.S., Adeishvili, N., Schneider, F., Watanabe, H., Tsunoda, S.P., Vogt, A., Elstner, M., Oertner, T.G., and Hegemann, P. (2014). Conversion of channelrhodopsin into a light-gated chloride channel. Science. 344, 409-412.
Selected Publications–Development of Optogenetics
Boyden, E.S., Zhang, F., Bamberg, E., Nagel, G., and Deisseroth, K. (2005). Millisecond- timescale, genetically targeted optical control of neural activity. Nat. Neurosci. 8, 1263- 1268.
Zhang, F., Wang, L.-P., Brauner, M., Liewald, J.F., Kay, K., Watzke, N., Wood, P.G., Bamberg, E., Nagel, G., Gottschalk, A., and Deisseroth, K. (2007). Multimodal fast optical interrogation of neural circuitry. Nature. 446, 633-639.
Aravanis, A.M., Wang, L.-P., Zhang, F., Meltzer, L.A., Mogri, M.Z., Schneider, M.B., and Deisseroth, K. (2007). An optical neural interface: in vivo control of rodent motor cortex with integrated fiberoptic and optogenetic technology. J. Neural. Eng. doi: 10.1088/1741- 2560/4/3/S02
Gradinaru, V. Mogri, M., Thompson, K.R., Henderson, J.M., and Deisseroth, K. (2009). Optical deconstruction of Parkinsonian neural circuitry. Science. 324, 354-359.
Berndt, A., Lee, S.Y., Wietek, J., Ramakrishnan, C., Steinberg, E.E., Rashid, A.J., Kim, H., Park, S., Santoro, A., Frankland, P.W., Iyer, S.M., Pak, S., Åhrlund-Richter, S., Delp, S.L., Malenka, R.C., Josselyn, S.A., Carlén, M., Hegemann, P., and Deisseroth, K. (2016). Structural foundations of optogenetics: determinants of channelrhodopsin ion selectivity. Proc. Natl. Acad. Sci. USA. 113, 822-829.
Allen, W.E., Chen, M.Z., Pichamoorthy, N., Tien, R.H, Pachitariu, M., Luo, L., and Deisseroth, K. (2019). Thirst regulates motivated behavior through modulation of brainwide neural population dynamics. Science. doi: 10.1126/science.aav3932
Perspectives
Zhang, F. Vierock, J., Yizhar, O., Fenno, L.E., Tsunoda, S., Kianianmomeni, A., Prigge, M., Berndt, A., Cushman, J., Polle, J., Magnuson, J., Hegemann, P., and Deisseroth, K. (2011). The microbial opsin family of optogenetic tools. Cell. 147, 1446-1457.
Grote, M., Engelhard, M., and Hegemann, P. (2014). Of ion pumps, sensors and channels–Perspectives on microbial rhodopsins between science and history. Biochim. Biophys. Acta. 1837, 533-545.
Deisseroth, K., and Hegemann, P. (2017). The form and function of channelrhodopsin. Science. 357. doi: 10.1126/science.aan5544
Fenno, L., Yizhar, O., and Deisseroth, K. (2011). The development and application of optogenetics. Annu. Rev. Neurosci. 34, 389-412.
Deisseroth, K. (2015). Optogenetics: 10 years of microbial opsins in neuroscience. Nat. Neurosci. 18, 1213-1225.

The Surprise Element: A Distinctive Characteristic of Creative Scientists, Artists, and Comedians
Truly creative works of science and art produce unexpected and surprising results—just like the punch line of a good joke that generates an unfamiliar twist on a familiar idea.
Acceptance remarks
Acceptance remarks, Karl Deisseroth
I am tremendously honored to accept this award along with my friends and colleagues Peter Hegemann and Dieter Oesterhelt. Ours has been a story of international scientific cooperation and deep friendship over many years, from our elucidation of the microbial transduction of light into ion flux, to illuminating the mysteries of emotion, in health and in psychiatric disease.
In the same sense that congestive heart failure, the weakening of a pump, is accompanied by a variety of symptoms, we must ask: What are, materially, the diverse features of debilitating psychiatric disease? Due to our limited understanding, these disorders continue to cause worldwide morbidity and mortality comparable to, or greater than, that of any other illness. While Covid-19 reshapes epidemiology, the pandemic has brought greater urgency to our search for understanding in psychiatry; the trauma and intense stress experienced across the globe have intensified symptom expression in mental illness, and most of my new patients these days frame the acute exacerbations of their suffering in terms of the pandemic and its consequences.
This hope for physical understanding did not only bring me to the field of psychiatry, but sustained me during personally difficult years of residency training, and throughout the past twenty years of caring for patients with mental illness. This aspiration also guided my laboratory to develop an approach for mapping individual features of adaptive and maladaptive brain states onto causal and precisely defined elements of neural circuitry.
My earliest formal scientific education provided insight into the capability of high-resolution mechanistic studies to reveal deep mysteries of biology. Isolating the position of individual atoms in microbial light-activated membrane proteins and elucidating their precise roles allowed us to explore the operation of biological systems as large and complex as the mammalian brain—and in so doing, to illuminate the cellular-activity basis for survival drives such as thirst and hunger, the intricate cellular dynamics involved in sensation and perception, and even complex cognitive processes such as social interaction.
Coming full circle over the past twenty years, we now can approach causal and material understanding of psychiatric disease-related states such as anxiety and dissociation, at the level of highly-resolved physical components. This story exemplifies the power of taking a molecular approach to explore complex biological systems–even for the disorders that define psychiatry. The hope brought to psychiatry by detailed studies of microbial membrane proteins is an uplifting story for all to share.
The scope of this fifty-year story also exemplifies how science proceeds in unpredictable ways: here, from a solvent-induced color change noticed by Dieter Oesterhelt while prepping lysed archaea, to the study of light responses in single-celled green algae by Peter Hegemann, to our structural and mechanistic studies together of microbial light-gated ion channels, and finally to our study of complex internal representations and behaviors by recruiting individually-specified cells with light. In these difficult times but with hope for the future grounded in science, I am deeply grateful to our lab members and collaborators over the years, and to the global community. The process of science truly brings the many faces of humanity together.
Acceptance remarks, Peter Hegemann
I am extremely grateful to receive the 2021 Lasker Award. To share the prize with my early mentor Dieter Oesterhelt and my long-standing friend and collaborator Karl Deisseroth is a special source of joy.
Dieter’s discovery of bacteriorhodopsin fifty years ago as a light driven proton pump is a milestone in photoreceptor research. For many decades bacteriorhodopsin served as a model that helped spur countless advances. It also became a template for further investigation of light-sensitive microbial proteins.
My group studied the photoreceptor molecules of Chlamydomonas and other algae over many years until we understood the function of most of the amino acids that make up the tiny biological photodiodes called channelrhodopsins. The wide repertoire of biophysical and biochemical techniques built upon the foundation of earlier research was instrumental in helping gain deeper knowledge of these remarkable proteins.
In a great collaboration with Karl, we engineered the channelrhodopsins with the hope of better understanding Nature’s concepts. Ultimately, we were able to make microbial opsins available for neuroscience applications, creating a field now called optogenetics. Karl is to be commended for transferring the knowledge about channelrhodopsins to the neuroscience community.
Progress in science to some extent depends upon risky decisions of individuals. At times the results of these decisions can only be appreciated later in one’s career. We all rely upon generous and enthusiastic coworkers and upon international collaborators who suffered and enjoyed with us the ups and downs of often painstaking work. Together, we shared both the patience and the joy that comes from working in wild unexplored territories in order to bring discoveries about tiny proteins found in bacteria and algae to unexpected applications.
I am grateful for the opportunity that this Award gives us to look back to the discovery decades ago of the first microbial rhodopsin, all the way up to the wide use of channelrhodopsin in the neurosciences, and then forward toward the promise of clinical applications of optogenetics.
I look forward to celebrating together with my co-laureates Dieter and Karl, with our coworkers and families at home, and with members of the Lasker Jury.
Acceptance remarks, Dieter Oesterhelt
Being honored with this Award brings back the times more than fifty years ago when I was a young biochemist searching to expand my competency, but not yet having found a topic of my own. I knew that this was not easy because it had to be something new, in my phantasy something unheard of, but where to find it?
It is always helpful to keep the eyes open and to remain curious. In my case it was the dramatic color change of the so-called purple membrane of Halobacterium salinarum upon lipid extraction. This observation could not be explained by standard chemistry. The rest of the story fits to the dictum of my mentor the Nobel Prize-winning biochemist Feodor Lynen: “Be naive, make an experiment!” And the consequences of this observation and the following experiments became an obsession not only for me and later my lab, but for quite some time a multitude of other laboratories. Now, after more than fifty years, scientific progress with bacteriorhodopsin as a model for membrane proteins reached another peak by the creation of a new field for application of microbial retinal proteins: optogenetics. Therefore, it makes me very happy to share the award with Karl Deisseroth, and the discoverer of channelrhodopsin, Peter Hegemann, my former student.
A large number of colleagues, coworkers and PhD students have contributed to this success, which was not guaranteed at the beginning. On the contrary! In the first years after the discovery of bacteriorhodopsin, the scientific community was very skeptical about the claim of a light-driven proton pump. Skepticism was even stronger towards the claim of bacteriorhodopsin as a second route of photosynthesis in Nature. I remember very vividly the comment of Feodor Lynen when I told him about my idea and the first experiments demonstrating the existence of a proton pump: “I don’t believe it, but I wish you are right!”
Beside the expansion of research into bacteriorhodopsin illuminating its molecular mechanism in ever greater detail, two thematic extensions were essential in my lab: first, the search for more microbial retinal proteins, such as halorhodopsin, as well as a systems biological approach to understand their functional context in the cell. Second, a search for potential applications.
Another development turned out to be a real breakhrough: The discovery of channelrhodopsin by Peter Hegemann and its joint use with halorhodopsin by Karl Deisseroth, who promoted tremendously the new field of optogenetics within the neurosciences.
